Ogden, N. H., Bouchard, C., Brankston, G., Brown, E. M., Corrin, T., Dibernardo, A., Drebot, M. A., Fisman, D. N., Galanis, E., Greer, A., Jenkins, E., Kus, J. V., Leighton, P. A., Lindsay, L. R., Lowe, A.-M., Ludwig, A., Morris, S. K., Ng, V., Vrbova, L., Waddell, L., et Wood, H. (2022). Maladies infectieuses. Dans P. Berry et R. Schnitter (éd.), La santé des Canadiens et des Canadiennes dans un climat en changement : faire progresser nos connaissances pour agir. Ottawa (Ontario) : gouvernement du Canada.
Auteur principal
Nick H. Ogden (Agence de la santé publique du Canada )
Auteurs collaborateurs
Catherine Bouchard (Agence de la santé publique du Canada)
Gabrielle Brankston (Université de Guelph)
Elizabeth M. Brown (Santé publique Ontario)
Tricia Corrin (Agence de la santé publique du Canada)
Antonia Dibernardo (Agence de la santé publique du Canada)
Michael A. Drebot (Agence de la santé publique du Canada)
David N. Fisman (Université de Toronto)
Eleni Galanis (Centre de contrôle des maladies de la Colombie-Britannique et Université de la Colombie-Britannique)
Amy Greer (Université de Guelph)
Emily Jenkins (Université de la Saskatchewan)
Julianne V. Kus (Santé publique Ontario et Université de Toronto)
Patrick A. Leighton (Université de Montréal)
L. Robbin Lindsay (Agence de la santé publique du Canada)
Anne-Marie Lowe (Agence de la santé publique du Canada)
Antoinette Ludwig (Agence de la santé publique du Canada)
Shaun K. Morris (Hospital for Sick Children et Université de Toronto)
Victoria Ng (Agence de la santé publique du Canada)
Linda Vrbova (Agence de la santé publique du Canada)
Lisa Waddell (Agence de la santé publique du Canada)
Heidi Wood (Agence de la santé publique du Canada)
Les changements climatiques ont une incidence sur le risque de maladies infectieuses. Selon les données probantes, l’émergence récente de la maladie de Lyme au Canada a été favorisée par le réchauffement du climat, ce qui a rendu une plus grande partie du pays propice aux espèces de tiques qui sont porteuses de cette maladie. L’émergence d’autres maladies transmises par des insectes, comme l’encéphalite équine de l’Est, a possiblement été favorisée par le réchauffement du climat. En outre, des épidémies d’infections imputables au virus du Nil occidental ont probablement été favorisées par la variabilité des conditions météorologiques et climatiques, qui va augmenter en raison des changements climatiques. On sait également que des risques découlant d’un très grand éventail d’autres maladies infectieuses peuvent être fonction des conditions météorologiques et climatiques. L’évolution des tendances géographiques et saisonnières de ces maladies en Amérique du Nord de même que le risque accru d’importation de maladies sensibles au climat en provenance de contrées plus lointaines, sont susceptibles de poser des risques accrus pour la population canadienne au cours des prochaines décennies. Les mesures d’adaptation comprennent des évaluations des risques et des vulnérabilités, des systèmes intégrés de surveillance et d’alerte précoce qui exploitent des technologies émergentes, et l’approche « Un monde, une santé » (aussi connu sous le nom d’Une seule santé) qui englobe la santé humaine, animale et environnementale.
Messages clés
En raison des changements climatiques, bon nombre de maladies considérées comme étant « sensibles au climat » sont plus susceptibles d’émerger ou de réémerger au Canada et dans le monde entier. Ces maladies comprennent celles transmises par des arthropodes vecteurs (p. ex., virus du Nil occidental, maladie de Lyme), celles transmises directement de l’animal à l’humain (zoonoses, p. ex., rage, syndrome cardio-pulmonaire à hantavirus), celles transmises directement d’un humain à un autre (p. ex., grippe saisonnière, infections entérovirales) et celles pouvant être contractées par inhalation à partir de sources environnementales (p. ex., infection à Cryptococcus, maladie du légionnaire).
Des maladies infectieuses nouvelles au Canada peuvent se propager vers le nord à partir des États-Unis, et d’ailleurs dans le monde, par des personnes, des marchandises ou des animaux sauvages. Les effets socioéconomiques indirects des changements climatiques peuvent avoir une incidence sur la capacité des pays à prévenir et à contrôler les maladies infectieuses à l’échelle mondiale, ce qui accroît la probabilité que de nouvelles maladies arrivent au Canada par le biais des déplacements et de la migration des humains.
On s’attend à ce que les changements climatiques rendent l’environnement canadien plus propice aux arthropodes vecteurs (comme les moustiques et les tiques) et à la transmission de nouvelles maladies infectieuses. Par exemple, des maladies transmises par le moustique déjà au Canada, comme le virus du Nil occidental, qui cause habituellement un nombre limité d’infections chaque année, peuvent produire des épidémies dans un climat plus variable connaissant des événements météorologiques extrêmes plus fréquents.
Les effets potentiels des changements climatiques sur les maladies infectieuses sont déterminés au moyen d’études de modélisation, tandis que la surveillance des maladies a permis de cerner des changements au chapitre de l’occurrence des maladies infectieuses et, dans certains cas, de lier ces changements aux effets récents des changements climatiques. Ces études se limitent en grande partie aux maladies que les humains contractent à partir d’arthropodes vecteurs (insectes et tiques) et directement des animaux.
Le Canada a une importante capacité d’adaptation qui lui permet de faire face aux maladies infectieuses, étant donné ses robustes mécanismes de surveillance et d’intervention nationaux en matière de santé publique qui se rattachent à des réseaux nationaux et internationaux, de son système de santé solide et de sa capacité à innover sur le plan technologique. Il est également un chef de file pour ce qui est des approches « Un monde, une santé » (aussi connu sous le nom d’Une seule santé) qui tiennent compte des facteurs humains, animaux et environnementaux combinés, mettant à profit des connaissances provenant d’une multitude de disciplines et de secteurs. De telles approches sont essentielles à la planification en prévision des maladies infectieuses émergentes et réémergentes, y compris celles liées aux changements climatiques.
Le Canada accroît également sa capacité à réagir aux effets des changements climatiques sur les maladies infectieuses. Cette capacité sera renforcée par les mégadonnées et les technologies génomiques modernes, l’observation de la Terre à partir de satellites et l’exploration Web, de même que par les approches axées sur la « science participative » pour ce qui est de la surveillance des impacts des changements climatiques sur les maladies infectieuses.
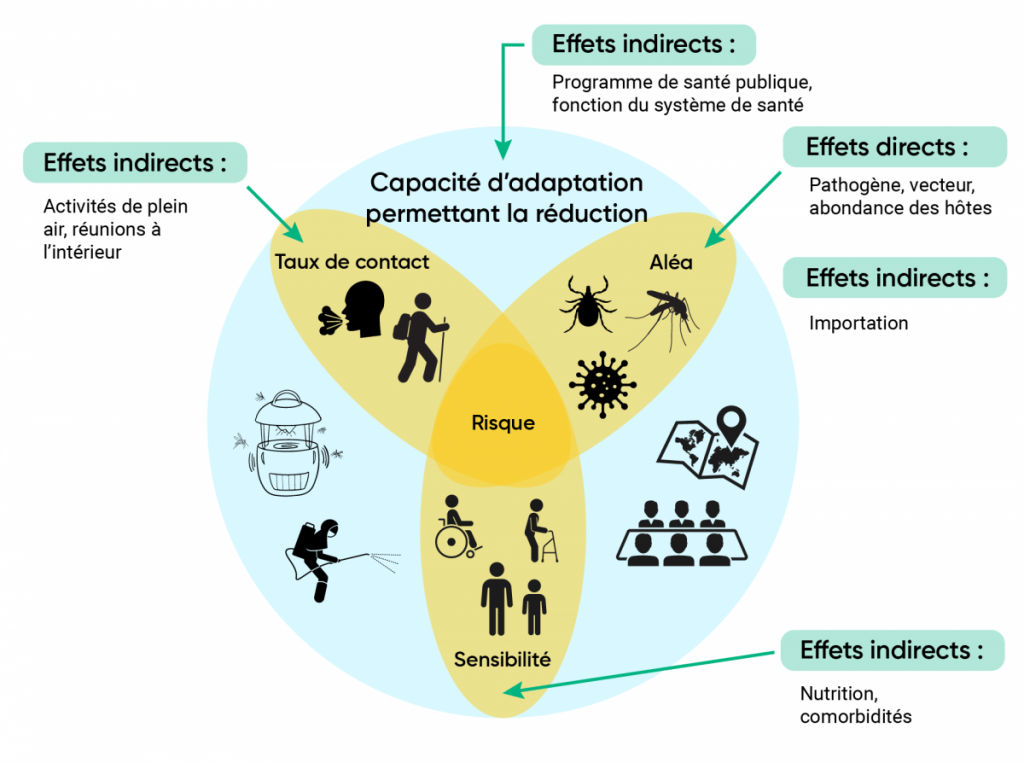
Éléments constitutifs de la vulnérabilité aux maladies infectieuses dans le contexte des changements climatiques.
Figure 6.1
Les trois éléments constitutifs du risque qui se recoupent sont les aléas et le taux de contact (qui déterminent ensemble l’exposition), et la sensibilité. L’adaptation (représentée par le cercle bleu en arrière-plan) est fonction de la capacité à atténuer ces trois éléments constitutifs du risque et à y réagir. Les flèches vertes illustrent les effets directs et indirects possibles des changements climatiques.
Aperçu des impacts des changements climatiques sur les maladies infectieuses
Catégorie d’impact et d’aléa sanitaire
Causes liées au climat
Effets possibles sur la santé
Maladies infectieuses transmises par des arthropodes vecteurs
Accroissement de l’abondance et de l’aire de répartition géographique des vecteurs que l’on retrouve au Canada en raison de l’accélération des taux de reproduction et d’un taux de survie plus élevé
Effets de la variabilité des conditions météorologiques et des événements météorologiques extrêmes sur les taux de reproduction et de survie des moustiques qui entraînent des changements rapides dans les populations
Accroissement de la probabilité d’établissement des vecteurs exotiques et des maladies qu’ils transmettent (agents pathogènes) en raison des taux de reproduction plus rapides et d’un taux de survie plus élevé des vecteurs exotiques une fois qu’ils sont transportés au Canada
Développement plus rapide des agents pathogènes chez les moustiques vecteurs
Incidence accrue des maladies à transmission vectorielle endémiques au Canada (p. ex., maladie de Lyme, infection par le virus du Nil occidental, encéphalite équine de l’Est)
Augmentation des épidémies de maladies transmises par les moustiques endémiques au Canada (infection par le virus du Nil occidental, encéphalite équine de l’Est, virus du sérogroupe de la Californie)
Propagation au Canada de maladies transmises par les tiques endémiques aux États-Unis (p. ex., ehrlichiose monocytique) et les moustiques (p. ex., infection par le virus La Crosse)
Risque accru de transmission autochtone de maladies tropicales/subtropicales transmises par les moustiques Aedes (p. ex., dengue, chikungunya, Zika)
Impacts sur les services de santé
Maladies infectieuses transmises directement par les animaux (maladies zoonotiques ou zoonoses)
Changements dans les taux de reproduction et de survie des hôtes naturels qui sont des animaux sauvages et d’autres espèces entraînant des changements dans les aires géographiques et les niveaux d’aléa pour les humains et les taux de contact avec ces derniers
Effets des conditions météorologiques sur les taux de reproduction et la survie d’espèces comme les rongeurs qui peuvent entraîner des changements rapides pour ce qui est de leur abondance
Changements (augmentation à certains endroits, diminution à d’autres) au chapitre des tendances géographiques et temporelles du risque de zoonoses transmises directement (p. ex., rage, brucellose)
Possibilité d’accroissement de la fréquence des éclosions de certaines zoonoses transmises par les rongeurs (p. ex., syndrome pulmonaire à hantavirus)
Impacts sur les services de santé
Maladies infectieuses contractées par inhalation à partir de sources environnementales
Accroissement possible du taux de survie et de reproduction des champignons dans les sols et d’autres environnements terrestres
Accroissement possible de la prolifération des bactéries Legionella en raison de l’utilisation accrue des climatiseurs
Augmentation de l’incidence des infections fongiques en suspension dans l’air ou en aérosol et variation de leur portée géographique (p. ex., cryptococcose)
Augmentation des éclosions de légionellose
Impacts sur les services de santé
Maladies infectieuses émergentes
Accroissement du risque d’émergence, de propagation aux humains et de transmission au Canada en raison des modifications découlant des changements climatiques en lien avec l’écologie des zoonoses à l’échelle internationale
Accroissement de la propagation mondiale des zoonoses qui sont également transmissibles de personne à personne en raison de l’augmentation des voyages internationaux, y compris la migration des populations humaines
Probabilité et fréquence accrues d’épidémies et de pandémies (p. ex., SRAS et COVID-19), y compris les maladies infectieuses d’origine hydrique, alimentaire, zoonotique et à transmission vectorielle
Transmission accrue d’agents pathogènes et de maladies infectieuses pouvant non seulement avoir des impacts sur la santé physique, mais sur la santé mentale, spirituelle et psychologique
Impacts sur les services de santé
Liste d’acronymes
ADMC analyse décisionnelle multicritères
EEV encéphalomyélite équine du Venezuela
EJ encéphalite japonaise
FJ fièvre jaune
FPMR fièvre pourprée des montagnes Rocheuses
FVR fièvre de la vallée du Rift
GOARN Réseau mondial d’alerte et d’action en cas d’épidémie
MCM modèle climatique mondial
MMPB maladie mains-pieds-bouche
OMS Organisation mondiale de la Santé
PIE période d’incubation extrinsèque
RCP profils représentatifs d’évolution de concentration
RMISP Réseau mondial d’information en santé publique
RSI Règlement sanitaire international
VEEE virus de l’encéphalite équine de l’Est
VELC virus de l’encéphalite de La Crosse
VESL virus de l’encéphalite de Saint-Louis
VJC virus de Jamestown Canyon
VNO virus du Nil occidental
VSGC virus du sérogroupe de la Californie
VSSH virus snowshoe hare
VUSU virus Usutu
WGS séquençage du génome entier
6.1
Introduction
Partager
Les maladies infectieuses continuent d’émerger et de réémerger dans le monde entier, comme l’illustrent les épidémies et les pandémies de maladies nouvelles et existantes, telles que la maladie à coronavirus 2019 (COVID-19) causée par le coronavirus SRAS-CoV-2, le virus Ebola en Afrique, le coronavirus du syndrome respiratoire du Moyen-Orient (CoV-SRMO) dans le Moyen-Orient, et les infections par les virus Zika et chikungunya, la fièvre jaune et le virus de la dengue dans les Amériques. De nombreuses maladies infectieuses sont sensibles au climat; par conséquent, les changements climatiques peuvent avoir un effet direct ou indirect sur la transmission de maladies. Au Canada, on s’attend à ce que les risques actuels de maladies infectieuses augmentent et que de nouvelles maladies apparaissent à mesure que le climat se réchauffe, ce qui est préoccupant pour la santé humaine. Le présent chapitre examine les impacts des changements climatiques sur les risques de maladies infectieuses importantes pour la santé publique au Canada. Il donne suite aux évaluations nationales antérieures sur les changements climatiques et la santé afin de déterminer l’état des connaissances sur les risques actuels et prévus liés aux maladies infectieuses, d’évaluer la vulnérabilité et de définir les efforts en matière de santé publique qui s’imposent pour protéger la population canadienne. Les maladies infectieuses liées aux impacts des changements climatiques sur la transmission des agents pathogènes dans l’eau et les aliments sont examinées au chapitre 7 : Qualité, quantité et sécurité de l’eau, et au chapitre 8 : Salubrité et sécurité des aliments, respectivement.
La composition démographique de la population canadienne évolue, avec le vieillissement de la population (Statistique Canada, 2016) et le nombre accru de personnes atteintes de maladies chroniques, deux facteurs pouvant modifier la sensibilité des gens aux aléas des maladies infectieuses (Dye, 2014). Dans le présent chapitre, on explique que le risque comporte deux composantes principales, soit la probabilité d’exposition et la sensibilité (c.-à-d. la gravité du résultat), tandis que la vulnérabilité de la population est considérée comme le risque jumelé à la capacité d’adaptation (figure 6.1). Cela s’assimile à d’autres versions dans la littérature portant sur le lien entre le risque et la vulnérabilité, où la vulnérabilité est une composante du risque qui combine la sensibilité, l’exposition et la capacité d’adaptation de la population humaine (GIEC, 2014). La probabilité d’exposition dépend du niveau d’aléa, c’est-à-dire du nombre d’organismes infectieux (c.-à-d. humains, micro-organismes, arthropodes vecteurs, hôtes animaux infectieux) dans l’environnement d’un individu, et du taux de contact des personnes non infectées avec l’aléa (figure 6.1). Dans le contexte des maladies infectieuses, la capacité d’adaptation s’entend de la capacité des systèmes de santé publique à repérer, à prévenir et à contrôler les maladies, et des systèmes de santé à réduire au minimum l’impact de la maladie par un traitement rapide et efficace. Les changements climatiques peuvent avoir une incidence sur chacun des éléments constitutifs de la vulnérabilité par des effets directs sur l’existence et le niveau d’aléa, par des effets indirects sur les taux de contact avec l’aléa, par une augmentation ou une diminution de la sensibilité de la population et par une incidence sur la capacité d’adaptation en tant que telle (figure 6.1).
Éléments constitutifs de la vulnérabilité aux maladies infectieuses dans le contexte des changements climatiques.
Figure 6.1
Les trois éléments constitutifs du risque qui se recoupent sont les aléas et le taux de contact (qui déterminent ensemble l’exposition), et la sensibilité. L’adaptation (représentée par le cercle bleu en arrière-plan) est fonction de la capacité à atténuer ces trois éléments constitutifs du risque et à y réagir. Les flèches vertes illustrent les effets directs et indirects possibles des changements climatiques.
6.1.1
Émergence et réémergence des maladies infectieuses
Partager
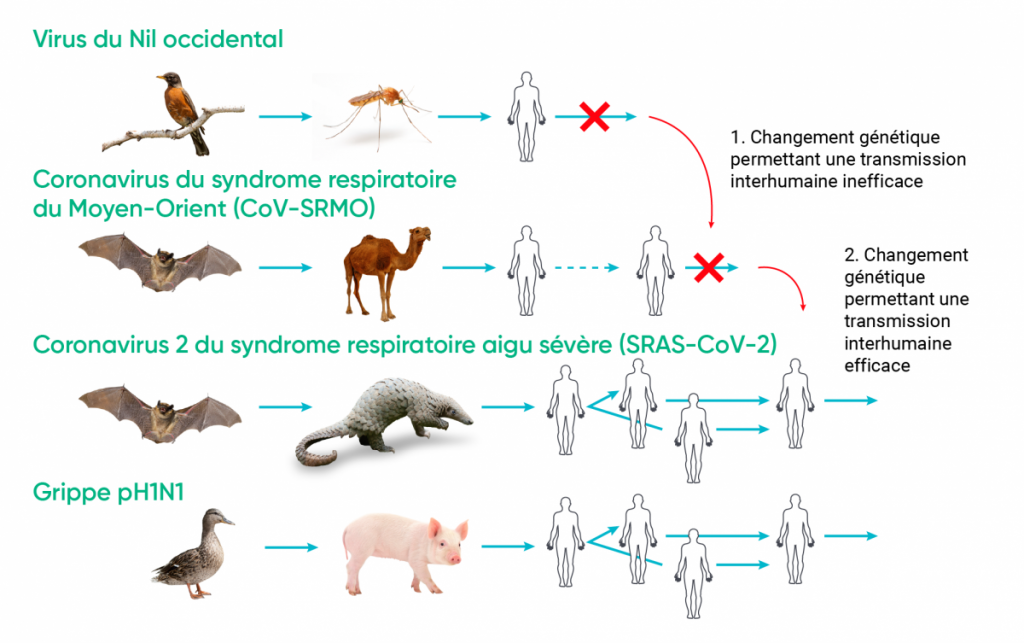
Les maladies infectieuses émergent à la suite de changements dans leur aire géographique, de la « propagation interespèce » et de « l’émergence adaptative ». Il y a propagation interespèce lorsque certains changements environnementaux ou socioéconomiques permettent à un agent zoopathogène déjà transmissible à l’humain d’entrer en contact avec celui-ci (p. ex., virus Nipah). L’émergence adaptative est imputable à un changement génétique dans un micro-organisme infectant les animaux, habituellement des espèces sauvages, de sorte qu’il devient transmissible à l’humain (c.-à-d. qu’il devient une zoonose) et possiblement transmissible entre les humains (p. ex., SARS-CoV) (Ogden et coll., 2017).
Il y a de multiples facteurs d’émergence des maladies, notamment ceux qui sont associés à la mondialisation et à l’environnement. Ces derniers comprennent notamment les changements climatiques, les changements sociaux et démographiques, et les changements sur le plan des systèmes et des politiques de santé publique (Semenza et coll., 2016). Ces mêmes facteurs peuvent être responsables de la réémergence des maladies endémiques (c.-à-d. l’accroissement de leur incidence ou leur réapparition sous forme d’épidémies). Les changements climatiques peuvent avoir une incidence directe sur l’émergence et la réémergence des maladies infectieuses en raison des effets sur la survie des agents pathogènes, des taux de survie et de reproduction des arthropodes vecteurs (p. ex., moustiques, tiques, puces) et, dans le cas des zoonoses, des effets sur l’abondance des hôtes animaux. Ces facteurs déterminent le potentiel de propagation d’un agent pathogène chez les humains ou, dans le cas des zoonoses, chez les hôtes animaux; ce potentiel est représenté par le taux de reproduction de base R0. Si R0 est égal ou supérieur à un (ce qui signifie qu’une infection entraîne au moins une autre infection), l’agent pathogène peut persister et se propager, mais s’il est inférieur à un, il disparaîtra (Anderson et May, 1991). Les effets directs des changements climatiques qui provoquent l’émergence de la maladie à un endroit particulier sont en fait ceux qui font passer la valeur R0 de moins de un ou un et plus à cet endroit.
Les changements climatiques peuvent avoir des impacts indirects sur l’émergence et la réémergence des maladies, en influant sur d’autres changements environnementaux et sociaux et en réduisant la capacité d’intervention des systèmes de santé publique (p. ex., les événements météorologiques extrêmes peuvent perturber la capacité de la santé publique à contrôler les éclosions de maladies). Les effets des changements climatiques sur les écosystèmes, y compris sur la biodiversité, peuvent modifier les aléas posés par les zoonoses par des effets complexes sur les communautés d’espèces sauvages (Altizer et coll., 2013; Cable et coll., 2017). D’autres changements susceptibles d’avoir une incidence sur l’émergence et la réémergence peuvent être liés aux initiatives de santé publique en matière d’adaptation qui visent à réduire les risques plus généraux pour la santé liés aux changements climatiques. Par exemple, les efforts visant à réduire les îlots de chaleur dans les zones urbaines par le virage écologique des villes (Beaudoin et Gosselin, 2016) et les mesures de gestion des inondations (voir le chapitre 3 : Aléas naturels) peuvent accroître les risques de zoonoses causées par la faune et l’incidence des maladies à transmission vectorielle (Medlock et Vaux, 2011; Millins et coll., 2017). L’utilisation accrue de la climatisation pour lutter contre la chaleur urbaine pourrait accroître les risques de légionellose (Fitzhenry et coll., 2017).
Les changements climatiques peuvent avoir des impacts négatifs sur les économies, particulièrement celles des pays à revenu faible ou intermédiaire, qui pourraient directement ou indirectement, en raison d’une fréquence accrue des conflits, nuire à la lutte contre les maladies infectieuses et contribuer à l’intensification de la densité des agents infectieux (Ogden, 2017). Les impacts économiques et les déplacements de populations peuvent simultanément entraîner une augmentation de la migration économique ou de l’arrivée de réfugiés, et donc l’importation de maladies infectieuses au Canada en provenance de l’étranger (Ogden, 2017). En outre, si les systèmes de santé ne sont pas résilients face aux changements climatiques (p. ex., résilients aux pannes d’électricité et de systèmes de communication associées aux événements météorologiques extrêmes; voir le chapitre 10 : Adaptation et résilience des systèmes de santé), les changements climatiques pourraient avoir un impact sur la capacité de repérer les maladies infectieuses émergentes ou réémergentes et d’y réagir (Mayhew et Hanefeld, 2014; Ebi et coll., 2018; Commission mondiale sur l’adaptation, 2019). La gamme de changements climatiques prévus comprend les changements à long terme dans les régimes de température et de précipitations, l’accroissement de la variabilité du climat et la fréquence accrue des événements météorologiques extrêmes, qui variera selon la région géographique du Canada (Bush et Lemmen, 2019). Ces changements auront une incidence directe et indirecte sur différents risques de maladies infectieuses de façon idiosyncratique (Ogden et Lindsay, 2016).
6.1.2
Maladies infectieuses et évaluations antérieures des changements climatiques et de la santé
Partager
Depuis 2008, il y a eu six évaluations nationales, régionales ou internationales des impacts des changements climatiques sur les risques de maladies infectieuses et la vulnérabilité à cet égard (tableau 6.1). Deux évaluations canadiennes ont été réalisées : une portait sur la santé, en 2008 (Charron et coll., 2008), et l’autre, sur la santé dans un rapport plus général du gouvernement du Canada sur les impacts des changements climatiques et l’adaptation à ces changements, en 2014 (Berry et coll., 2014a). Les évaluations internationales pertinentes pour le Canada comprenaient des chapitres sur la santé et sur l’Amérique du Nord du Groupe d’experts intergouvernemental sur l’évolution du climat (GIEC) (Romero-Lankao et coll., 2014; Smith et coll., 2014) et des évaluations nationales menées aux États-Unis (Beard et coll., 2016; Ebi et coll., 2018).
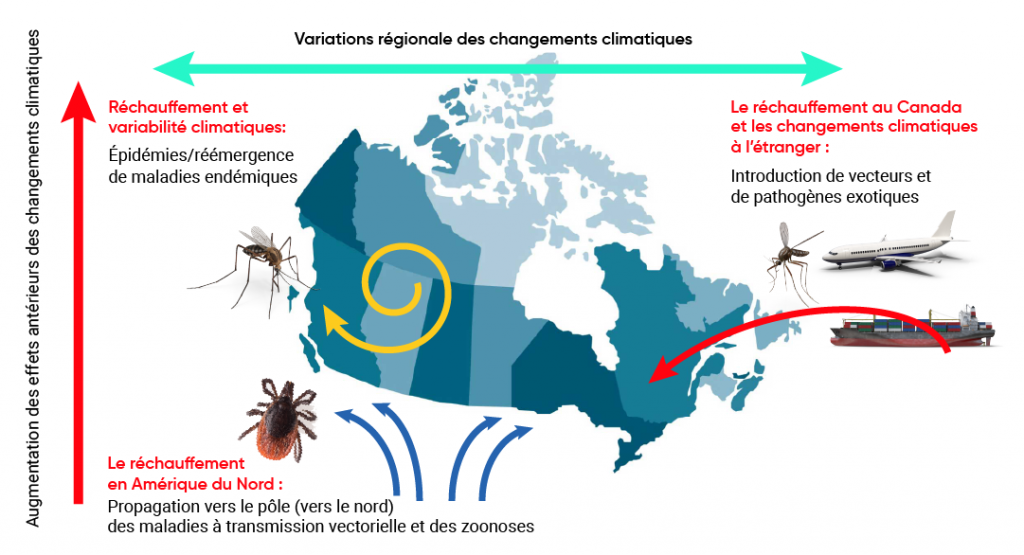
Toutes les évaluations ont déterminé que les maladies transmises par les arthropodes (tels que les moustiques, les tiques et les puces) sont les plus sensibles au climat, et que celui-ci détermine leur occurrence et leur abondance par ses effets sur la survie et la reproduction des vecteurs et sur le développement d’agents pathogènes dans certains vecteurs. Les évaluations semblent indiquer que le réchauffement climatique est susceptible d’accroître les risques associés à ces maladies, mais la plupart ont indiqué une confiance modérée dans la capacité de déterminer les maladies à transmission vectorielle qui seront touchées et l’ampleur des effets. Cela s’explique par la complexité des cycles de transmission des maladies à transmission vectorielle. De multiples déterminants non climatiques, dont les changements environnementaux et ceux liés à l’aménagement du territoire, en particulier pour les zoonoses à transmission vectorielle comme le virus du Nil occidental (VNO) et la maladie de Lyme, et les mesures de contrôle, surtout pour les maladies exotiques transmises par les moustiques comme le paludisme et la dengue, sont importants pour déterminer l’occurrence et l’abondance des vecteurs et des agents pathogènes. Des évaluations antérieures ont mis en évidence l’incidence du climat sur la sensibilité aux aléas endémiques au Canada et aux États-Unis, y compris la peste et les infections à hantavirus transmise par des rongeurs, la maladie de Lyme et d’autres maladies transmises par les tiques, comme la fièvre pourprée des montagnes Rocheuses (FPMR), et les maladies transmises par les moustiques, comme le VNO, le virus de l’encéphalite équine de l’Est (VEEE) et les virus du sérogroupe de la Californie. Les évaluations ont révélé qu’il est fort probable que les changements climatiques modifient les aires géographiques et la durée des saisons de transmission (Charron et coll., 2008; Berry et coll., 2014a; Smith et coll., 2014; Beard et coll., 2016; Ebi et coll., 2018). La figure 6.2 illustre les voies par lesquelles les changements climatiques peuvent avoir une incidence sur les risques de maladies infectieuses au Canada, selon des évaluations antérieures.
Effets des changements climatiques sur les risques de maladies infectieuses au Canada.
Source
Ogden et Gachon, 2019.
L’évaluation canadienne de 2008 a fourni des projections fondées sur un modèle de la propagation de la maladie de Lyme vers le nord, c’est-à-dire des États-Unis au Canada, en raison de l’expansion de l’aire de répartition du vecteur de la tique Ixodes scapularis causée par les changements climatiques (Charron et coll., 2008). La propagation de la tique le long des trajectoires déterminées par le climat et l’émergence de la maladie de Lyme au Canada ont depuis été documentées (Berry et coll., 2014a; Smith et coll., 2014; Beard et coll., 2016; Ebi et coll., 2018) (tableau 6.1). Ces mêmes évaluations ont également permis d’évaluer les risques d’introduction de maladies exotiques à transmission vectorielle, telles que le paludisme, la dengue et le virus chikungunya, associés aux changements climatiques. Les températures à la hausse devraient accroître l’étendue des zones géographiques de l’Amérique du Nord propices à la transmission d’agents pathogènes par des vecteurs exotiques et à la survie des vecteurs de maladie des moustiques exotiques (p. ex., espèces d’Aedes).
Tableau 6.1
Résumé des constatations des évaluations relatives aux effets des changements climatiques sur les maladies infectieuses, à l’exclusion des maladies d’origine hydrique et alimentaire
Évaluation
Maladies infectieuses sensibles au climat décelées
Impacts anticipés des changements climatiques
Données probantes concernant les impacts des changements climatiques sur les risques de maladie
Adaptation et capacité d’adaptation au moment de l’évaluation
Chapitre 5 : Répercussions des changements climatiques sur les maladies transmises par l’eau, les aliments, les vecteurs et les rongeurs dans Santé et changements climatiques : Évaluation des vulnérabilités et de la capacité d’adaptation au Canada (Charron et coll., 2008)
Maladies transmises par les tiques
Maladies transmises par les moustiques
Hantavirus
Peste
Propagation des maladies transmises par les tiques et les moustiques vers le nord, c’est-à-dire des États-Unis vers le Canada
Importation de maladies exotiques transmises par les moustiques (p. ex., dengue, paludisme)
Solide capacité d’adaptation du Canada en matière d’évaluation des risques, de surveillance, de prévention et de contrôle, mais lacunes relevées au chapitre des connaissances sur l’écologie des maladies, les effets du climat, la capacité spécialisée, la surveillance et les systèmes d’avertissement
Chapitre 7 : Santé humaine dans Vivre avec les changements climatiques au Canada : perspectives des secteurs relatives aux impacts et à l’adaptation(Berry et coll., 2014a)
Voir ci-dessus
Voir ci-dessus
Données probantes sur l’émergence de la maladie de Lyme le long de trajectoires déterminées par les facteurs climatiques, ainsi que sur l’incidence croissante des cas chez l’humain
Voir ci-dessus
Chapitre 11 : Santé humaine : Incidences, adaptation et avantages accessoires dans le cinquième Rapport d’évaluation du Groupe d’experts intergouvernemental sur l’évolution du climat (GIEC) (Smith et coll., 2014)
Maladies transmises par les moustiques, telles que le paludisme et la dengue, dans les pays à revenu faible ou intermédiaire
Maladies transmises par les tiques en Europe et en Amérique du Nord
Peste en Asie et en Amérique du Nord
Hantavirus en Amérique du Nord
Augmentation possible de l’incidence et de l’expansion de l’aire de répartition des maladies transmises par les moustiques et les tiques
Aucune
Besoin d’aborder les risques et la capacité d’adaptation dans tous les pays par les moyens suivants :
Réduction de la pauvreté
Amélioration de la nutrition ainsi que des services de santé publique et de santé de base
Cartographie des vulnérabilités
Mise sur pied de systèmes d’alerte précoce liés aux programmes de lutte
Chapitre 26 : Amérique du Nord dans le cinquième Rapport d’évaluation du GIEC(Romero-Lankao et coll., 2014)
Maladies transmises par les moustiques, comme le virus du Nil occidental
Maladies transmises par les tiques, en particulier la maladie de Lyme
Augmentation possible de l’incidence et de l’expansion de l’aire de répartition des maladies transmises par les moustiques et les tiques
Risque de propagation de maladies exotiques transmise par les moustiques
Données probantes sur l’émergence de la maladie de Lyme le long de trajectoires déterminées par les facteurs climatiques au Canada
Nécessité d’améliorer les ensembles de données et les modèles pour comprendre les effets des changements environnementaux par rapport aux autres déterminants du risque de maladies à transmission vectorielle et des systèmes d’alerte précoce
Chapitre 5 : Vector-Borne Diseases dans The Impacts of Climate Change on Human Health in the United States: A Scientific Assessment (en anglais seulement) (Beard et coll., 2016)
Maladies transmises par les moustiques comme le virus du Nil occidental et la dengue
Maladies transmises par les tiques comme la maladie de Lyme
Peste
Variation de l’aire de répartition géographique, de la saisonnalité et de la période d’activité (début et durée) du vecteur
Aucune
Besoin de modèles améliorés pour comprendre et prévoir les effets des changements météorologiques et climatiques par rapport à d’autres déterminants du risque de maladies à transmission vectorielle, et d’observations sur le terrain à l’appui
Chapitre 14 : Human Health dans Impacts, Risks, and Adaptation in the United States: Fourth National Climate Assessment (Ebi et coll., 2018; en anglais seulement)
Maladies transmises par les moustiques comme le virus du Nil occidental et la dengue
Maladies transmises par les tiques comme la maladie de Lyme
Changements au chapitre des aires géographiques, de la répartition saisonnière et de l’abondance des vecteurs de maladies
Risques accrus de maladies transmises par les moustiques à l’étranger, qui peuvent se propager sur le territoire américain
Aucune
Nécessité de réaliser des évaluations de la vulnérabilité et de l’adaptation, d’élaborer des plans d’intervention, d’assurer une surveillance intégrée des maladies à transmission vectorielle et mettre en place des systèmes d’alerte précoce, comme la prévision des maladies
6.1.3
Gestion des risques de maladies infectieuses
Partager
L’adaptation aux maladies infectieuses liées au climat passe d’abord par l’évaluation des risques pour la santé et la détermination des populations plus vulnérables à leurs impacts. Il convient d’évaluer la capacité des systèmes de santé publique à déceler les maladies infectieuses émergentes et réémergentes en exerçant surveillance, et de prévenir et contrôler celles-ci grâce à la promotion de la santé ou la prise de mesures plus proactives (p. ex., au point de vaccins), de même que la capacité des systèmes de soins de santé à réduire au minimum les conséquences des maladies infectieuses. Les évaluations fournissent des renseignements sur la vulnérabilité aux maladies infectieuses et déterminent les mesures les plus efficaces pour la réduire (Berry, 2008). De façon générale, des évaluations antérieures ont indiqué qu’en Amérique du Nord, la capacité des systèmes de santé publique d’évaluer les risques liés aux maladies infectieuses émergentes et réémergentes et de les déceler, de les prévenir et de les contrôler est robuste (voir le tableau 6.1). Le risque pour la plupart des Canadiens et des Canadiennes est faible en raison du statut socioéconomique relativement élevé d’une grande proportion de la population qui évolue dans un milieu qui limite de nombreux risques de maladie, et en raison de la robustesse des infrastructures et des systèmes de soins de santé et de santé publique. En outre, des progrès ont été réalisés pour combler les lacunes en matière de connaissances, de surveillance, de capacité et de système d’alerte précoce relevées dans les évaluations précédentes (voir la section 6.4 Adaptation en vue de réduire les risques pour la santé).
6.2
Méthodes et approche
Partager
Le présent chapitre analyse les impacts des changements climatiques sur les risques que présentent pour les humains les maladies infectieuses importantes pour la santé publique au Canada, à l’exception des maladies infectieuses transmises dans l’eau potable et celle utilisée à des fins récréatives et dans les aliments, qui sont traitées dans des chapitres distincts (voir le chapitre 7 : Qualité, quantité et sécurité de l’eau et le chapitre 8 : Salubrité et sécurité des aliments). Le chapitre est un compte rendu narratif rédigé par des experts en la matière. Toutefois, afin d’appuyer l’équipe de rédaction, un examen rapide a été mené pour répertorier la majeure partie de la littérature nationale et internationale sur les impacts des conditions météorologiques, du climat et des changements climatiques sur les maladies infectieuses. L’examen portait sur les cinq domaines examinés dans le chapitre répertoriés ci-dessous.
Maladies à transmission vectorielle :
Maladies exotiques transmises par les moustiques, y compris celles pour lesquelles les humains sont les principaux hôtes (p. ex., paludisme, dengue)
Maladies transmises par les moustiques endémiques au Canada (p. ex., maladie du VNO)
Maladies transmises par des insectes autres que les moustiques (p. ex., peste)
Maladies transmises par les tiques (p. ex., maladie de Lyme)
Maladies infectieuses transmises directement de personne à personne (p. ex., influenza, infections entérovirales)
Maladies infectieuses transmises par inhalation de sources environnementales (p. ex., cryptococcose, légionellose)
Capacité du Canada de s’adapter à l’évolution des risques liés aux maladies infectieuses
L’examen rapide a suivi le cadre général des examens de la portée proposé d’abord par Arksey et O’Malley (2005), puis peaufiné au cours des 15 dernières années (Levac et coll., 2010; Peters et coll., 2015; Tricco et coll., 2016). Il a recensé et caractérisé toutes les recherches internationales disponibles sur les changements climatiques et les maladies infectieuses à l’aide d’une méthodologie systématique et reproductible. Un protocole a été élaboré au préalable et définit la portée de l’examen rapide, la stratégie de recherche exhaustive et tous les outils utilisés pour trier les citations et extraire l’information de la documentation (disponible sur demande). La recherche électronique a été effectuée au moyen d’Embase, de PubMed et de Global Health en septembre 2018 afin de recenser la documentation pertinente publiée en anglais et en français sur les maladies infectieuses qui examinait également les effets des conditions météorologiques, du climat et des changements climatiques. Une recherche dans la littérature grise a été effectuée manuellement dans divers sites Web gouvernementaux et scientifiques afin de repérer les rapports qui n’étaient pas indexés dans les bases de données électroniques. Les doublons ont été supprimés dans le logiciel de gestion des références EndNote (EndNote X7, Clarivate Analytics), et les citations uniques ont été téléchargées dans le logiciel de revue systématique en ligne DistillerSR (DistillerSR, Evidence Partners, Ottawa, Canada). La sélection de la pertinence a été effectuée par deux examinateurs, travaillant indépendamment, à l’aide d’un outil de sélection de la pertinence mis au point au préalable. Toutes les citations qui ont été incluses après l’examen de la pertinence ont été obtenues, et leur pertinence a été confirmée. La recherche a ensuite été caractérisée à l’aide de l’outil de caractérisation des données conçu. Des publications plus récentes ont été recensées au cours du processus d’examen des chapitres.
Lorsqu’il existe suffisamment de renseignements, le présent chapitre établit le niveau de confiance à l’égard des orientations et de l’intensité des effets des changements climatiques. La détermination de la sensibilité des maladies aux conditions climatiques ou météorologiques semblerait fournir des données probantes limitées sur les effets des changements climatiques; le jumelage de ce type de données aux projections des effets des changements climatiques semblerait produire des données probantes de qualité moyenne, tandis que les changements recensés à l’égard des risques liés aux maladies infectieuses imputables aux récents changements climatiques sembleraient fournir des données probantes solides relativement aux effets des changements climatiques.
6.3
Risques climatiques pour la santé, impacts prévus des changements climatiques et données probantes sur les impacts à ce jour
Partager
6.3.1
Maladies à transmission vectorielle
Partager
6.3.1.1 Effets des changements climatiques sur l’écologie et l’épidémiologie des vecteurs et sur la transmission des maladies à transmission vectorielle
Les cycles de vie de nombreux arthropodes vecteurs et les impacts des conditions météorologiques et climatiques sur ces cycles de vie ont été étudiés de manière intensive en laboratoire, sur le terrain et par des études de modélisation, et sont examinés ailleurs (Ogden et Lindsay, 2016). La façon dont les changements climatiques et météorologiques peuvent avoir une incidence sur les arthropodes vecteurs et la transmission des maladies à transmission vectorielles est résumée dans l’encadré 6.1. Bien que les effets des conditions météorologiques et climatiques soient les mêmes pour les arthropodes vecteurs, la façon dont ils touchent expressément les cycles de vie et les cycles de transmission est hautement idiosyncrasique parmi les différents vecteurs et agents pathogènes (figure 6.3). Les changements climatiques peuvent avoir une incidence sur le risque de maladies à transmission vectorielle en modifiant les facteurs de risque liés au comportement social comme la perception du risque et l’adoption de comportements préventifs (Bouchard et coll., 2018). La croissance démographique, les déplacements et des facteurs sociaux et économiques (p. ex., changement de l’exposition au virus de l’encéphalite à tiques en Europe de l’Est en raison de l’évolution de l’exposition de la population à la suite de l’effondrement du bloc communiste) ont également été associés à des taux différentiels d’exposition humaine aux vecteurs et aux maladies à transmission vectorielle (Randolph, 2004). La façon dont les changements climatiques peuvent avoir une incidence indirecte sur les risques en raison de ces facteurs doit faire l’objet d’une étude plus approfondie.
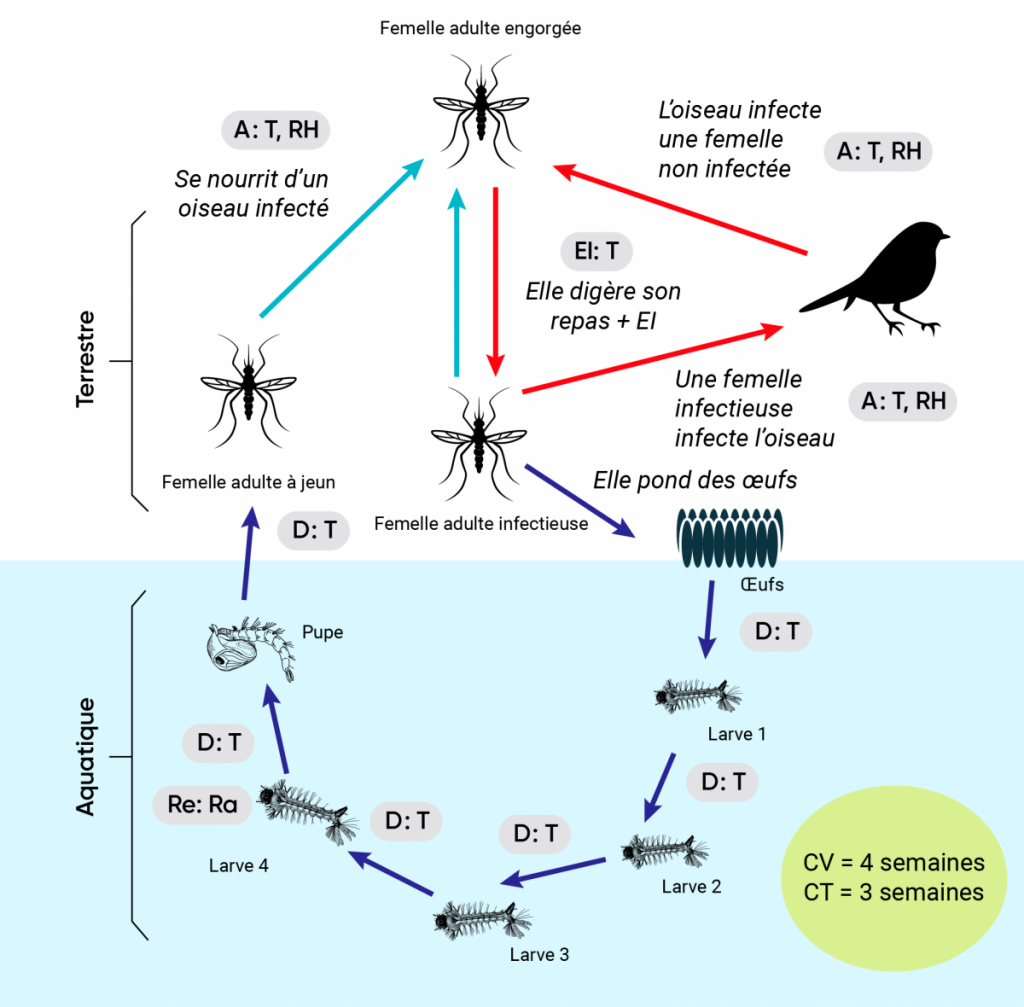
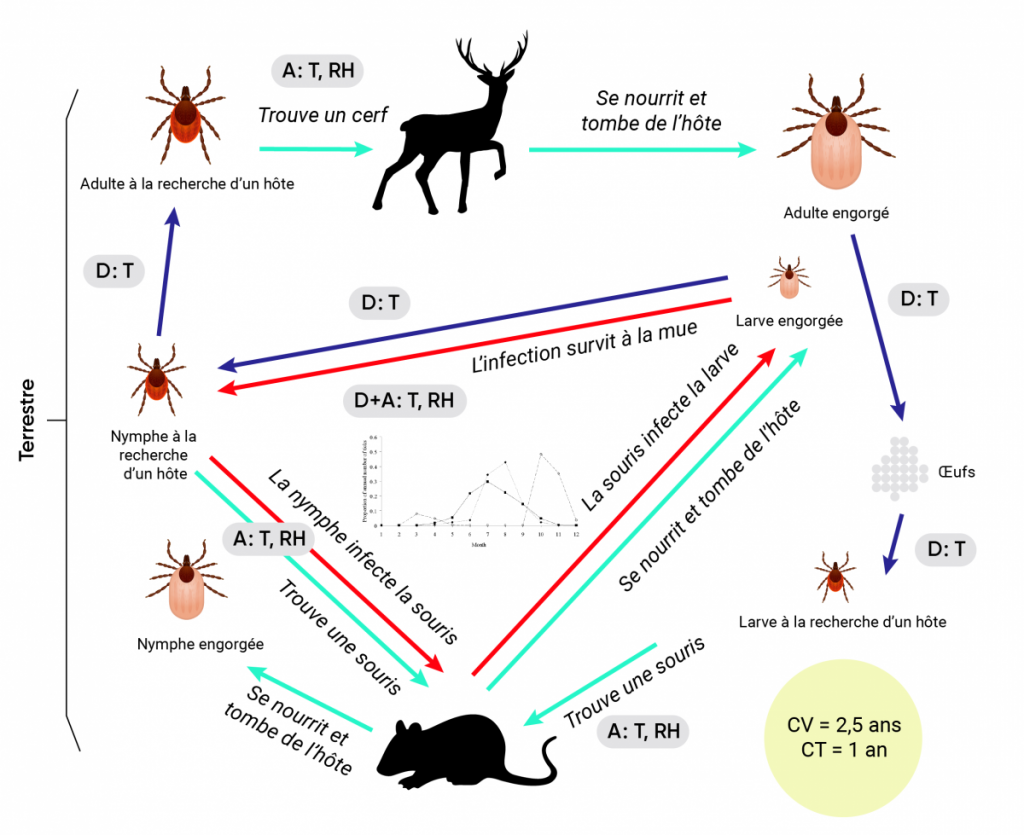
Figure 6.3
Partager
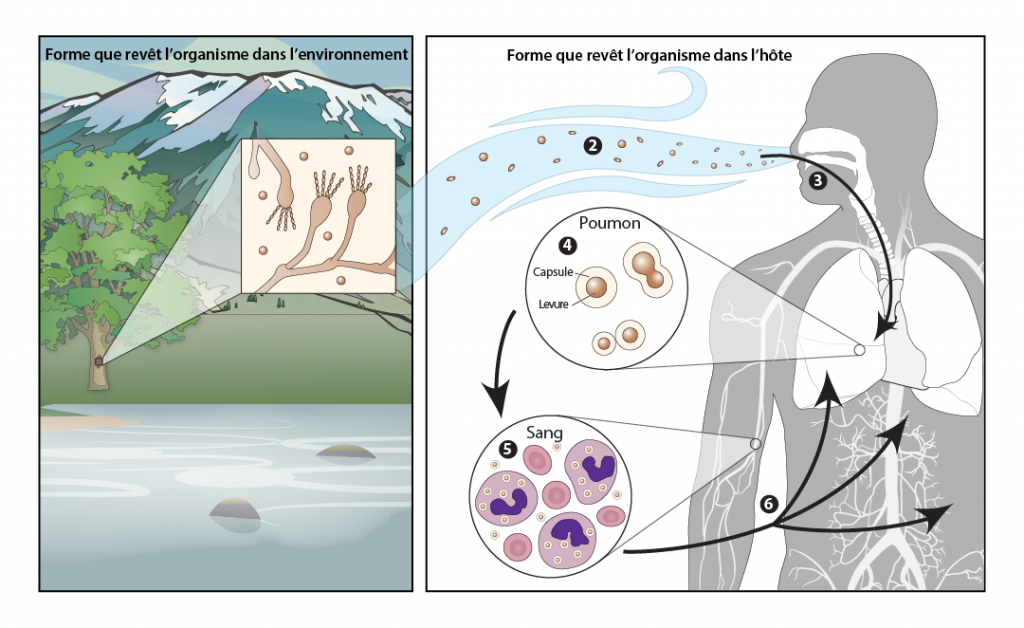
Impacts des conditions météorologiques et climatiques sur le cycle de vie des diptères et des tiques ixodides vecteurs.
1 / 2
Figure 6.3
Les impacts des changements climatiques sont illustrés en utilisant dans le diagramme A l’exemple du cycle de vie du moustique Culex pipiens et de son rôle dans la transmission du virus du Nil occidental (VNO). Le cycle de vie de la tique Ixodes scapularis et son rôle dans la transmission de Borrelia burgdorferi, l’agent de la maladie de Lyme, est illustré dans le diagramme B. Dans les deux cas, les flèches bleu foncé et bleu pâle indiquent respectivement le développement et la recherche d’hôtes ou le détachement des hôtes, tandis que les flèches rouges indiquent les cycles de transmission des agents pathogènes. Les étapes auxquelles les conditions météorologiques et le climat (et, potentiellement, les changements climatiques) peuvent avoir une incidence sur le cycle de vie (CV) du vecteur et le cycle de transmission (CT) des agents pathogènes sont indiquées par les cases grisées dans lesquelles « A » correspond aux effets sur l’activité, « D » correspond aux effets sur les taux de passage d’un stade à l’autre, « EI » correspond aux effets sur la période d’incubation extrinsèque, « Re » correspond aux effets sur la reproduction, « T » correspond aux effets de la température, « Ra » correspond aux effets des précipitations et « RH » correspond aux effets de l’humidité. Au centre du cycle de transmission de B. burgdorferi (B) se trouve une illustration de l’activité saisonnière des tiques vectrices qui pourrait être touchée par le climat et les changements climatiques en raison des effets sur le développement et l’activité décrits dans le texte.
Source
Ogden et Lindsay, 2016.
6.3.1.2 Maladies exotiques transmises par les moustiques
Il y a un vaste éventail de maladies transmises par les moustiques qui ont une incidence sur la santé publique à l’échelle mondiale. Certaines, comme le paludisme, la dengue, la fièvre jaune et le chikungunya, qui sont transmis de personne à personne par les moustiques, figurent parmi les plus grandes menaces à la santé publique au monde, avec des millions de cas et de décès chaque année (OMS, 2019a). D’autres maladies, dont bon nombre sont des zoonoses transmises par les moustiques comme celles causées par et les virus de l’encéphalite équine du Sindbis et du Venezuela, surviennent de façon plus sporadique (tableau 6.2). Les maladies exotiques transmises par les moustiques ne sont pas établies au Canada en raison de nombreux facteurs, dont le climat. Dans le cas de la plupart des maladies exotiques transmises par des moustiques, le climat du Canada est actuellement trop froid pour les vecteurs ou pour le développement des agents pathogènes chez le moustique (Ng et coll., 2019). D’autres obstacles existent, notamment l’absence d’hôtes animaux pour certaines maladies (comme les primates pour le virus de la fièvre jaune), les normes en matière de logement, notamment les portes et les fenêtres qui empêchent l’entrée des vecteurs et les systèmes de climatisation, qui inhibent la transmission d’une personne à une autre par les moustiques (Reiter, 2001). Les barrières physiques doivent être surmontées pour que les agents pathogènes et les vecteurs soient transportés ici depuis l’étranger.
L’expansion de la dispersion géographique des maladies exotiques à transmission vectorielle au Canada exige l’introduction de vecteurs et d’agents pathogènes exotiques en provenance des pays où ils sont endémiques. On s’attend à ce que l’augmentation du commerce et des déplacements à l’échelle mondiale facilite la migration internationale des moustiques vecteurs (infectés ou non) et des voyageurs infectés, accroissant ainsi la possibilité d’introduire des vecteurs et des agents pathogènes dans des régions où ils étaient absents auparavant (Tatem et coll., 2006; Tatem et coll., 2012; Semenza et coll., 2016). Il y a trois scénarios d’introduction d’agents pathogènes exotiques à transmission vectorielle au Canada (encadré 6.2).
L’impact des changements climatiques sur la santé publique variera selon le scénario d’émergence et pourrait accroître la possibilité que les Canadiens et les Canadiennes contractent une infection lors d’un voyage à l’étranger, que la transmission autochtone soit de courte durée (c.-à-d. locale, confinée au Canada) lorsque des vecteurs compétents sont déjà établis au Canada, et qu’il y ait endémicité permanente de nouvelles maladies (Ng et coll., 2019).
Les vecteurs de maladies exotiques transmises par les moustiques sont sensibles aux conditions météorologiques, lesquelles devraient être touchées par les changements climatiques. Des expériences sur le terrain et en laboratoire démontrent la sensibilité à la température des moustiques des espèces Aedes et Anopheles, qui sont les principaux vecteurs des maladies exotiques les plus graves à l’échelle mondiale (Brady et coll., 2013; Shapiro et coll., 2017). En général, des températures plus chaudes, un taux d’humidité élevé et une augmentation des précipitations facilitent le cycle de vie des moustiques en soutenant le développement et la survie des larves et en prolongeant la durée de vie des adultes (Reeves et coll., 1994; Jetten et Focks, 1997; Paaijmans et coll., 2009; Yang et coll., 2009). Ces conditions climatiques influeront sur la transmission des agents pathogènes de la façon suivante :
Réduction du temps de développement des œufs chez les moustiques femelles adultes, diminuant ainsi le temps entre les repas de sang et accroissant leur fréquence
Raccourcissement de la période d’incubation extrinsèque (PIE), ce qui permet aux moustiques de devenir infectieux plus rapidement (Davis, 1932; Reeves et coll., 1994; Jetten et Focks, 1997; Paaijmans et coll., 2009; Paaijmans et coll., 2013; Xiao et coll., 2014)
Augmentation de la longévité des moustiques, permettant aux moustiques infectieux de piquer un plus grand nombre de personnes (Yang et coll., 2009)
Toutefois, bon nombre de ces relations dépendantes des conditions météorologiques sont complexes; les relations peuvent être non linéaires, peuvent avoir des effets opposés selon les circonstances et peuvent être influencées par des facteurs non climatiques (encadré 6.1).
La mesure dans laquelle les changements climatiques auront ou ont déjà eu un effet sur la répartition mondiale de nombreux moustiques et maladies transmises par les moustiques est incertaine. En effet, la relation entre le climat et les maladies à transmission vectorielle, en particulier celles transmises de personne à personne par les moustiques, est complexe, et la répartition spatiotemporelle des vecteurs et des agents pathogènes qu’ils transportent dépend d’une série de facteurs qui ne sont pas directement liés au climat. Ces facteurs comprennent : la mobilité accrue et l’interconnectivité des personnes et des biens (Junxiong et Yee-Sin, 2015; Semenza et coll., 2016; Tabachnick, 2016; Lindsey et coll., 2018; Romeo-Aznar et coll., 2018); l’urbanisation et d’autres changements dans l’aménagement du territoire (Junxiong et Yee-Sin, 2015; Jones et O’Neill, 2016; Semenza et coll., 2016; Asad et Carpenter, 2018; Romeo-Aznar et coll., 2018); les facteurs socioéconomiques (KC et Lutz, 2017; Reina Ortiz et coll., 2017; Lindsey et coll., 2018; Moreno-Madriñán et Turell, 2018; Romeo-Aznar et coll., 2018); les changements démographiques, y compris ceux imputables à l’immigration et à la croissance démographique (Asad et Carpenter, 2018; Lindsey et coll., 2018; Romeo-Aznar et coll., 2018); l’immunité de la population (Anyamba et coll., 2012; Larrieu et coll., 2014; Semenza et coll., 2016); l’évolution et l’adaptation génétiques (Gubler, 2007; Tsetsarkin et coll., 2007; Tabachnick, 2016); l’accès aux soins de santé (Reiter, 2008; Ooi et Gubler, 2009; Semenza et coll., 2016); et les programmes de lutte et d’intervention contre les vecteurs (Githeko et coll., 2000; Junxiong et Yee-Sin, 2015; Tasanee et coll., 2015). Les changements climatiques n’ont aucune incidence sur certains de ces facteurs, mais peuvent avoir un effet sur d’autres et, par le fait même, avoir des répercussions indirectes sur les maladies transmises par les moustiques.
Malgré l’incertitude, il est probable que les changements climatiques auront des effets sur la propagation des moustiques et des maladies transmises par les moustiques (confiance élevée), et il y aura des variations régionales partout dans le monde (Smith et coll., 2014). En raccourcissant la durée du cycle de vie, en prolongeant la survie des moustiques et en améliorant la transmission des agents pathogènes, les changements climatiques favoriseront l’expansion des populations de moustiques et d’agents pathogènes transmis par les moustiques à certains endroits et élargiront la répartition géographique de leurs niches écologiques dans de nombreux cas, principalement en direction du pôle et en altitude (Campbell et coll., 2015; Kraemer et coll., 2015; Samy et coll., 2016; Hertig, 2019; Kamal et coll., 2019; Kraemer et coll., 2019). Bien que les populations de moustiques puissent étendre leur territoire à de nouvelles régions géographiques, elles peuvent disparaître dans d’autres régions (Machado-Machado, 2012; Escobar et coll., 2016; Williams et coll., 2016).
On s’attend à ce que le réchauffement climatique augmente l’abondance des populations d’espèces de moustiques endémiques au Canada (Hongoh et coll., 2012) et améliore les conditions de transmission des agents pathogènes, ce qui créera un environnement plus propice à l’émergence de nouvelles maladies transmises par les moustiques selon les scénarios 1 et 2, surtout dans le sud du Canada (encadré 6.2). De plus, les conditions de température dans certaines parties du sud du Canada devraient devenir de plus en plus favorables pour les populations de moustiques Ae. aegypti et Ae. albopictus (moustique tigre), qui sont des vecteurs de la dengue, du chikungunya, du Zika et de la fièvre jaune. Les changements climatiques devraient faire en sorte que les températures se prêtent davantage à la transmission de virus (Ng et coll., 2017), accroissant ainsi la possibilité que l’émergence de maladies transmises par les moustiques selon le scénario 3 (encadré 6.2) se produise dans le sud du Canada. On anticipe que les conditions climatiques de la côte sud de la Colombie-Britannique ainsi que du Québec, du Nouveau-Brunswick, de la Nouvelle-Écosse et du sud de l’Ontario, deviennent adaptées sur le plan climatique pour l’espèce Ae. albopictus (Ogden et coll., 2014a). Les conditions climatiques de la côte sud de la Colombie-Britannique pourraient également devenir adaptées pour l’espèce Ae. aegypti (Campbell et coll., 2015; Kamal et coll., 2019). En 2016, un spécimen d’Ae. aegypti adulte a été collecté dans le sud de l’Ontario. Un très petit nombre d’adultes et de larves ont été trouvés au cours de l’année suivante dans le cadre du piégeage des moustiques et d’une surveillance accrue sur le terrain dans la région, ce qui porte à croire que cette espèce n’est pas établie au Canada (Windsor-Essex County Health Unit, 2019a). Cependant, l’espèce Ae. albopictus a été observée de façon sporadique dans plusieurs endroits précis du sud de l’Ontario depuis 2005. Il y a une petite région du Canada où le moustique Ae. albopictus s’est établi entre 2017 et 2018. Toutefois, à ce jour, rien n’indique que ces moustiques sont porteurs d’agents pathogènes pouvant provoquer des maladies chez les humains (Windsor-Essex County Health Unit, 2019b).
La probabilité d’émergence de nouvelles maladies à transmission vectorielle au Canada, pour les scénarios d’émergence 2 et 3, dépendra du nombre d’événements d’introduction d’agents pathogènes ou de vecteurs, ainsi que de la mesure dans laquelle les changements climatiques font du Canada un environnement plus propice à leur établissement. Certaines formes de déplacements mondiaux peuvent être imputables aux changements climatiques, par exemple, les réfugiés climatiques (McMichael et coll., 2012) et les changements dans les habitudes de voyage (Organisation mondiale du tourisme et Programme des Nations Unies pour l’environnement, 2008). Les changements climatiques peuvent donc avoir trois impacts qui accroissent la probabilité d’émergence selon les scénarios 2 et 3 :
Conditions climatiques plus favorables à la transmission au Canada
Augmentation de l’abondance d’agents pathogènes exotiques et de leurs vecteurs dans les pays autres que le Canada
Introduction accrue d’agents pathogènes et de vecteurs
Même sans les effets des changements climatiques, l’interconnectivité mondiale augmente (Findlater et Bogoch, 2018) et les Canadiens et les Canadiennes sont des voyageurs enthousiastes (Statistique Canada, 2019). Les déplacements locaux et mondiaux continueront donc de présenter un risque croissant d’introduction au Canada d’agents pathogènes exotiques par l’intermédiaire de vecteurs importés et d’humains infectés. Les voies d’introduction peuvent toutefois être prévisibles. Dans le cas du virus de l’encéphalite japonaise (VEJ), le scénario le plus probable d’introduction aux États-Unis serait l’arrivée de moustiques femelles infectés par le VEJ à bord d’avions en provenance de l’est de la Chine (Oliveira et coll., 2018). L’introduction du VEJ suivrait probablement une voie similaire vers le Canada, la côte de la Colombie-Britannique étant peut-être le lieu d’entrée le plus probable en raison d’une forte connectivité avec l’Asie sur le plan des voyages et du commerce. Cette région jouit également d’un climat tempéré et les réservoirs et moustiques vecteurs endémiques qui pourraient soutenir la transmission locale du VEJ, s’il est introduit, y sont établis.
Outre un climat propice, d’autres facteurs tels que la pauvreté et le manque d’accès aux infrastructures, aux soins de santé et aux mesures de lutte contre les maladies sont également importants pour la transmission endémique des agents pathogènes exotiques transmis de personne à personne par les moustiques (Ebi et coll., 2006a; Halstead, 2008). Les conditions sociales de la majeure partie de la population canadienne (en particulier la densité démographique et les conditions de logement) devraient limiter les taux de piqûres de moustiques, et les services de santé retirent les personnes infectées de la chaîne de transmission, ce qui fait qu’il est plus difficile pour ces agents pathogènes exotiques d’établir des cycles de transmission efficaces et soutenus (Berrang-Ford et coll., 2009; Ng et coll. 2019). Il est plus probable que l’impact sur la santé publique se fera sentir dans des cas autochtones occasionnels chez des Canadiens et des Canadiennes qui n’ont pas voyagé. Ceci pourrait avoir de graves conséquences pour la santé parce que le diagnostic peut être retardé en l’absence d’antécédents de voyage (Berrang-Ford et coll., 2009; Ng et coll., 2019). Un diagnostic tardif de cas « inattendus » de maladies exotiques transmises par les moustiques chez la population canadienne pourrait déboucher sur quelques chaînes de transmission et des éclosions limitées. Parmi les exemples provenant de l’extérieur du Canada, mentionnons l’introduction du virus du chikungunya en Europe méditerranéenne en 2007 et de la dengue en France, en Croatie et à Madère entre 2010 et 2013. Ces événements pourraient avoir été imputables à des conditions météorologiques particulièrement chaudes (Rezza et coll., 2007; Tomasello et Schlagenhauf, 2013).
En revanche, si les zoonoses exotiques transmises par les moustiques sont introduites, elles risquent davantage de devenir endémiques si les conditions environnementales, y compris le climat, sont favorables, car la transmission d’agents pathogènes parmi les hôtes réservoirs chez les animaux sauvages ou les animaux d’élevage ne serait pas grandement limitée, comme ce fut le cas pour le VNO. L’expansion de l’aire de répartition du virus Sindbis (VSIN) et du virus Usutu (VUSU), à l’extérieur de l’Afrique et en Europe, a fortement ressemblé à celle du VNO en Amérique du Nord (Weissenböck et coll., 2002; Ashraf et coll., 2015). Étant donné que les moustiques vecteurs et les réservoirs du VUSU et du VSIN sont déjà présents au Canada et que le climat dans les régions envahies de l’Europe est très semblable à celui des régions du sud du Canada, l’émergence du VSIN ou du VUSU en tant que virus endémiques est possible dans ce pays. Les infections par le VSIN et le VUSU produisent des infections désagréables, mais relativement bénignes chez les humains immunocompétents.
Une autre zoonose exotique transmise par les moustiques, le virus de la fièvre de la vallée Rift (VFVR), peut causer des infections graves et mortelles (OMS, 2019b), de sorte que la possibilité d’émergence de ce virus au Canada est une menace plus préoccupante. Il a été démontré que les espèces de moustiques endémiques au Canada sont des vecteurs compétents pour le VFVR, alors qu’un large éventail d’espèces sauvages et de bétail domestiqué sont des réservoirs animaux (encadré 6.2, tableau 6.2). On estime que le mécanisme d’introduction le plus probable en Amérique du Nord est celui des personnes infectées par le VFVR qui arrivent à bord d’un avion en provenance des zones endémiques touchées par une éclosion (Golnar et coll., 2014; Golnar et coll., 2018). Si le voyageur entre en contact avec des moustiques vecteurs endémiques, la transmission aux espèces sauvages et au bétail peut engendrer de l’endémicité.
Tableau 6.2
Principales maladies transmises par les moustiques à l’échelle mondiale et caractéristiques de leur transmission (vecteurs et hôtes), occurrence géographique et présence de vecteurs et d’hôtes au Canada et aux États-Unis
Maladie
Agent(s) pathogène(s)
Répartition géographique des cas chez l’humain
Endémicité aux États-Unis
Principaux vecteurs
Vecteurs établis au Canada?
S’ils sont absents du Canada, sont-ils établis aux États-Unis?
Principaux réservoirs
Principaux réservoirs présents au Canada?
Données indiquant une sensibilité au climat
Scénario d’émergence
Chikungunya
Virus Chikungunya
Afrique, Asie du Sud-Est, Philippines, îles du Pacifique, Moyen-Orient, Caraïbes, Amériques
Non endémique, mais des cas d’origine locale ont été signalés en 2014-2015 en Floride et au Texas
Aedes aegypti et Ae. albopictus
Population émergente d’Ae. albopictus dans une zone très limitée du sud de l’Ontario
Aedes aegypti dans les états du Sud jusqu’au sud de l’État de New York, Ae. albopictus dans les États du Sud et du Nord-Est, y compris ceux qui longent la frontière des provinces du centre et de l’est du Canada
Humains, primates sauvages (en Afrique)
Oui (humains), pas de primates sauvages
Oui
2
Dengue
Virus de la dengue, sérotypes 1 à 4
Asie, Pacifique, Amériques, Afrique, Caraïbes
Endémique à Porto Rico
Ae. aegypti et Ae. albopictus
Population émergente d’Ae. albopictus dans une zone très limitée du sud de l’Ontario
Aedes aegypti dans les états du Sud jusqu’au sud de l’État de New York, Ae. albopictus dans les États du Sud et du Nord-Est, y compris ceux qui longent la frontière des provinces du centre et de l’est du Canada
Humains, primates sauvages en Asie du Sud-Est et en Afrique de l’Ouest
Oui (humains), pas de primates sauvages
Oui
2
Encéphalite japonaise
Virus de l’encéphalite japonaise
Asie, îles du Pacifique, détroit de Torres d’Australie, Papouasie-Nouvelle-Guinée
Non endémique
Espèce de moustiques Culex
Cx. tarsalis dans l’ouest et le centre du Canada
Porcs domestiques et oiseaux sauvages
Oui
Oui
2
Encéphalite de La Crosse
Virus La Crosse
États-Unis (États du Haut-Midwest, du centre du littoral atlantique et du sud-est)
Endémique dans la plupart des États de l’Est, y compris les États qui longent la frontière du Canada
Aedes triseriatus
Aedes triseriatus dans l’est du Canada
Petits mammifères (tamia rayé et écureuil)
Oui
Oui
1
Malaria
Plasmodium spp.
Amérique centrale, Amérique du Sud, Afrique, Asie du Sud-Est, Moyen-Orient, sud-ouest du Pacifique
Non endémique
Moustiques Anophèles spp., en particulier An. gambiae et An. arabiensis
Anopheles quadrimaculatus dans l’est de l’Ontario et au Québec, Anopheles freeborni en Colombie-Britannique
Humains
Oui
Oui
2
Fièvre de la vallée du Rift
Fièvre de la vallée du Rift
Afrique orientale et australe, Arabie saoudite, océan Indien
Non endémique
Moustiques Aedes spp. qui se reproduisent dans les eaux de crue en Afrique
Certaines espèces d’Aedes établies au Canada peuvent transmettre le virus en laboratoire
Inconnus, peut-être le bétail, les équidés et les ongulés sauvages
Oui
Oui
3
Fièvre Sindbis
Virus Sindbis
Europe du Nord, Australie, Chine, Afrique du Sud
Non endémique
Moustiques ornithophiles Culex, Culiseta, Ochlerotatus et Aedes spp.
Culex pipiens et quelques Ochlerotatus et Aedes spp.
Oiseaux
Oui
Oui
2
Encéphalite de Saint-Louis
Virus de l’encéphalite de Saint-Louis
Centre, ouest et sud des États-Unis
Endémique dans les États du nord, de l’est et du centre, y compris les États qui longent la frontière du Canada
Culex tarsalis, C. pipiens, C. quinquefasciatus, C. nigripalpus
Culex tarsalis dans l’ouest et le centre du Canada et Culex pipiens le long de la frontière canado-américaine
Oiseaux
Oui
Oui
1
Virus Usutu
Virus Usutu
Afrique, Europe
Non endémique
Culex pipiens, Cx. neavei
Culex pipiens le long de la frontière canado-américaine
Oiseaux
Oui
Oui
2
Encéphalomyélite équine du Venezuela
Virus de l’encéphalite équine du Venezuela
Amérique du Sud et centrale, Mexique et cas jusqu’au sud des États-Unis
Non endémique
Moustiques Culex, Aedes, Mansonia, Psorophora, Deincerites et Ochlerotatus spp.
Oc. sollicitans dans l’est du Canada
Oc. taeniorhynchus le long de la majeure partie de la côte américaine et Culex melanoconion en Floride
Rongeurs, équidés
Oui
Oui
2
Fièvre jaune
Virus de la fièvre jaune
Afrique tropicale et subtropicale, Amérique du Sud
Non endémique
Ae. aegypti
Aedes aegypti dans les états au sud de l’État de New York
Humains, primates sauvages
Oui (humains), pas de primates sauvages
Oui
3
Zika
Virus Zika
Afrique, Asie du Sud-Est, Amérique du Sud
Non endémique, mais des cas acquis localement ont été signalés à Porto Rico, en Floride et au Texas de 2015 à 2016
Ae. aegypti, Ae. albopictus
Population émergente d’Ae. albopictus dans une zone très limitée du sud de l’Ontario
Aedes aegypti dans les états du Sud jusqu’au sud de l’État de New York, Ae. albopictus dans les États du Sud et du Nord-Est, y compris ceux qui longent la frontière des provinces du centre et de l’est du Canada
Humains, primates sauvages
Oui (humains), pas de primates sauvages
Oui
2
6.3.1.3 Maladies endémiques transmissibles par les moustiques au Canada
Les quatre arbovirus les plus importants sur le plan médical qui sont endémiques au Canada, soit le VNO, le VEEE, le virus de Jamestown Canyon (VJC) et le virus snowshoe hare (VSSH), sont transmis par des piqûres de moustiques femelles infectées. Les moustiques contractent les agents pathogènes d’hôtes réservoirs mammifères ou aviaires précis. Les principaux vecteurs du VNO sont Cx. pipiens et Cx. restuans dans l’est du Canada et Cx. tarsalis dans l’ouest (Kramer et coll., 2008), tandis que Culiseta melanura est le principal vecteur du VEEE (Armstrong et Andreadis, 2010). Les espèces de moustiques de catégories autres que Culex (p. ex., les espèces Aedes, Culiseta et Anopheles) sont les principaux vecteurs des virus du sérogroupe Californie (VSGC) comme le VSSH et le VJC (Drebot, 2015; Pastula et coll., 2015; Webster et coll., 2017). Pour ce qui est du VNO et du VEEE, un large éventail d’espèces d’oiseaux servent de réservoirs, y compris les corvidés et les passereaux (Kilpatrick et coll., 2006; Kramer et coll., 2008; Ludwig et coll., 2010; Reisen, 2013). Le principal réservoir du VJC est le cerf de Virginie (Andreadis et coll., 2008), tandis que les écureuils, les tamias et les lièvres sont les hôtes réservoirs du VSSH (Drebot, 2015). Un certain nombre de ces agents pathogènes peuvent aussi être transférés par transmission ovarienne, ce qui permet de moins dépendre des réservoirs des mammifères (Griot et coll., 1993).
D’autres agents viraux et bactériens transmis par les insectes sont également endémiques au Canada, mais ils sont moins actifs, ou leur présence est sous-étudiée. La prévalence du virus de l’encéphalite équine de l’Ouest semble avoir diminué au Canada au cours des dernières décennies, tandis que le virus de Cache Valley (VCV) a été responsable d’un certain nombre d’éclosions chez le bétail (c.-à-d. le mouton) en Ontario, au Québec et dans d’autres provinces, mais les cas d’infection chez l’humain sont probablement sous-déclarés (Drebot, 2015). Les arbovirus peuvent aussi parfois être transmis à la suite d’une transfusion sanguine ou d’une greffe de tissus (Fonseca et coll., 2005; Direction de la règlementation des agents pathogènes, 2010). Hormis cette possibilité, les humains sont des hôtes accidentels ou « terminaux » des arbovirus transmis par des moustiques, ce qui signifie que, bien qu’ils puissent être infectés, ils ne peuvent pas transmettre ces virus efficacement aux moustiques qui se nourrissent de leur sang en raison d’une virémie faible et transitoire ou d’une charge virale faible (Kramer et coll., 2008; Kulkarni et coll., 2015).
Environ 20 % des personnes exposées à des virus transmis par les moustiques, comme le VNO, le VEEE, le VJC ou le VSSH, développeront une maladie clinique aiguë dont les symptômes sont, notamment, la fièvre, des maux de tête, des éruptions cutanées, des nausées et des douleurs musculaires. La plupart des personnes infectées se rétablissent complètement, mais environ 1 % d’entre elles développent une maladie grave (p. ex., méningite, encéphalite, paralysie flasque aiguë et poliomyélite) causant des déficits neurologiques et cognitifs qui peuvent être prolongés ou permanents. Environ 10 % des cas graves sont mortels. La maladie peut frapper plus gravement les personnes de plus de 70 ans et celles qui ont des problèmes médicaux sous-jacents, comme l’obésité, le diabète, l’hypertension et les maladies du cœur. Cependant, le VSSH cause aussi des maladies neurologiques chez les enfants. Les personnes immunodéprimées sont aussi plus à risque (Petersen et coll., 2013a; Petersen et coll., 2013b; Sejvar, 2014; Badawi et coll., 2018). La gravité de la maladie varie et dépend du virus. Par exemple, le VEEE est l’une des maladies transmises par les moustiques les plus graves aux États-Unis, avec un taux de mortalité d’environ 33 % chez les personnes développant une maladie neurologique et des dommages cérébraux importants chez la plupart des survivants qui ont développé une maladie symptomatique (Centers for Disease Control and Prevention, 2018c). Le virus de l’encéphalite équine de l’Ouest et le VCV donnent lieu à une gamme similaire de symptômes — la majorité des cas sont asymptomatiques, mais, pour un pourcentage variable d’entre eux, l’infection se manifeste par une encéphalite, une méningite, une encéphalomyélite, une forte fièvre, une conscience altérée, une dysfonction neurologique, une méningite aseptique, une raideur de la nuque, des maux de tête, une myalgie, des tremblements, des nausées, des vomissements ou une infection des voies urinaires. Le taux de mortalité de l’encéphalite de Saint-Louis se situe entre 5 % et 20 %, mais on croit qu’il est beaucoup plus faible pour l’encéphalite équine de l’Ouest et l’infection par le VCV (Centers for Disease Control and Prevention, 2018d).
Les effets prévus des changements climatiques sur les maladies endémiques transmises par les moustiques au Canada sont les suivants : une expansion de l’aire de répartition vers le nord associée au réchauffement à long terme et une augmentation du comportement épidémique associée à la variabilité du climat et aux événements météorologiques extrêmes en raison de leurs effets sur la survie des vecteurs et les taux de reproduction (deux éléments qui influent sur l’abondance des vecteurs), les taux de morsure, la durée de la saison d’activité et la durée de la PIE. Les maladies endémiques transmises par les moustiques au Canada sont des zoonoses transmises par des animaux sauvages (oiseaux et mammifères). Les effets des changements climatiques sur les populations de ces animaux devraient avoir une incidence sur les cycles de transmission des agents pathogènes. Ces effets pourraient simplement entraîner une expansion de l’aire de répartition des hôtes vers le nord, mais des effets plus complexes sur la biodiversité des hôtes réservoirs peuvent se manifester. Par exemple, les changements au chapitre de l’abondance et de l’aire de répartition géographique des hôtes peuvent être limités par des conditions physiques (p. ex., obstacles au déplacement) et des processus biologiques (p. ex., réduction de l’accès aux aliments à des moments critiques du cycle de vie, comme les périodes de reproduction et d’élevage). Les changements qui en résultent dans la composition des espèces peuvent avoir des conséquences variées, comme des perturbations dans les relations prédateur-proie et hôte-parasite. Par conséquent, bien qu’il soit probable que la biodiversité des hôtes change en réponse aux nouvelles conditions climatiques, des incertitudes subsistent quant à la façon dont ces changements influeront sur le risque d’exposition des Canadiens et des Canadiennes aux maladies à transmission vectorielle (Varrin et coll., 2007). Selon toute vraisemblance, l’impact sera propre à l’écosystème ou à l’habitat, ce qui entraînera un ensemble incohérent d’augmentations et de diminutions de la biodiversité des populations hôtes, qui changera avec le temps à l’échelle du pays. Les événements météorologiques extrêmes causés par les changements climatiques, comme des sécheresses et des épisodes de chaleur, peuvent avoir des effets plus rapides sur les populations d’hôtes réservoirs en les amenant à chercher des sources d’eau dans les aires de reproduction des moustiques (Shaman et coll., 2005; Wang et coll., 2010; Harrigan et coll., 2014).
De nombreuses études de modélisation ont examiné la relation entre les variables climatiques (principalement la température et les précipitations) et l’infection par le VNO (humains, oiseaux ou moustiques infectés) au Canada (Wang et coll., 2011; Chen et coll., 2013; Tam et coll., 2014; Paz, 2015; Yoo et coll., 2016; DeFelice et coll., 2018). Toutefois, l’écologie du VEEE et des VSGC demeure sous-étudiée, probablement en raison des perceptions selon lesquelles ces virus ne sont pas aussi importants pour la santé publique, de même que du manque de données de surveillance détaillées. Cependant, on sait que la couverture terrestre, y compris la proximité et la taille de zones boisées de conifères et de terres humides, a une incidence sur la présence du VEEE et du VJC (Vander Kelen et coll., 2014; Rocheleau et coll., 2018) et qu’elle pourrait également être touchée par les changements climatiques.
L’impact des changements climatiques sur la transmission du VNO au Canada a fait l’objet de deux études qui sont parvenues aux mêmes conclusions. Chen et coll. (2013) ont examiné la transmission du VNO dans les Prairies, où Cx. tarsalis est le principal vecteur, et prévoient une prolongation de l’activité saisonnière de Cx. tarsalis infecté par le VNO, qui passerait de trois mois (c.-à-d. de juin à août) à cinq mois (c.-à-d. de mai à septembre) d’ici les années 2080. Les auteurs ont également prévu une expansion de l’aire de répartition vers le nord pour Cx. tarsalis et le VNO. Comme ce vecteur est également capable de transmettre le VCV, l’étendue et la prévalence de cette maladie peuvent également être touchées par cette expansion de Cx. tarsalis (Ayers et coll., 2018). Hongoh et coll. (2012) ont modélisé la répartition potentielle des populations de Cx. pipiens dans l’est du Canada en fonction des changements climatiques actuels et anticipés et prédisent une expansion semblable de l’aire de répartition de ce vecteur du VNO vers le nord.
Une meilleure compréhension de la façon dont les changements climatiques peuvent perturber les populations d’espèce réservoirs aviaires et mammifères permettrait une évaluation plus robuste des effets des changements climatiques, mais peu d’études ont réalisé ces évaluations en raison de données limitées et de contraintes méthodologiques. Les données actuelles indiquent que les aires de répartition (ou les enveloppes climatiques) dont le climat est adéquat pour de nombreuses espèces se déplaceront probablement vers le nord par suite du réchauffement des températures. Par exemple, les modèles de niche écologique pour 765 espèces portent à croire que les changements climatiques pourraient accroître la biodiversité dans le sud du Québec au cours de ce siècle à mesure que les espèces se déplacent vers le nord (Berteaux et coll., 2010; Chambers et coll., 2013). De même, de nombreuses espèces d’oiseaux qui nichent actuellement dans la partie nord de l’est des États-Unis sont susceptibles de se déplacer vers le nord au Canada, ce qui accroîtra la richesse des espèces d’oiseaux dans l’est du Canada (DesGranges et Morneau, 2010). La perte et la perturbation de l’habitat causées par les changements climatiques ou d’autres facteurs qui peuvent entraîner la fragmentation de l’habitat (Warren et Lemmen, 2014) peuvent avoir une incidence sur les populations des hôtes réservoirs aviaires et mammifères (Berteaux et Stenseth, 2006). La mesure dans laquelle ces effets positifs et négatifs sur les populations hôtes entraîneront une augmentation ou une réduction des risques de maladies transmises par les moustiques n’est pas encore claire et nécessite une étude plus approfondie (Salkeld et coll., 2013).
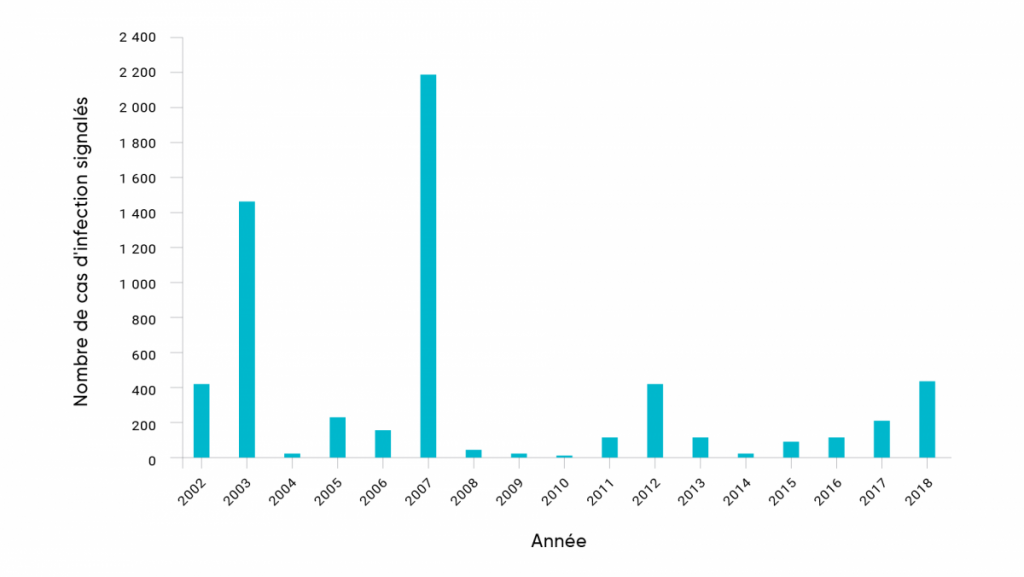
Les infections transmises par les moustiques sont signalées au Canada depuis de nombreuses décennies; toutefois, récemment, le nombre de cas d’infection à arbovirus semble augmenter (Ludwig et coll., 2019). Depuis 2002, le nombre de cas signalés chaque année de VNO chez l’humain, la seule maladie endémique transmise par les moustiques au Canada à déclaration obligatoire au pays, a fluctué considérablement au fil du temps à l’échelle nationale. Les cas signalés fluctuaient de 5 cas en 2010 à des sommets de 1 481 en 2003 (pendant l’invasion initiale au Canada) et de 2 215 cas en 2007, ce qui était associé à une abondance sans précédent de moustiques Cx. tarsalis dans les provinces des Prairies (figure 6.4). Cette augmentation pourrait être compatible avec les effets des conditions météorologiques et la variabilité du climat sur la dynamique du VNO (Ludwig et coll., 2019).
Nombre de cas de VNO déclarés chez l’humain chaque année au Canada.
Source
Gouvernement du Canada, 2019a.
Les variations géographiques au fil du temps ont également été très prononcées. En 2003 et 2007, la plupart des cas humains de VNO ont été signalés dans les Prairies (Alberta, Saskatchewan et Manitoba), mais en 2002, 2012 et 2018, la plupart des cas déclarés ont été détectés en Ontario et au Québec. Cette variabilité est, à tout le moins, partiellement compatible avec les effets de la variabilité des conditions météorologiques locales sur l’abondance des moustiques Cx. tarsalis, Cx. pipiens et Cx. restuans pendant les années d’éclosion, et une indication qu’une variabilité plus importante des conditions météorologiques causée par les changements climatiques pourrait engendrer un comportement épidémique accru des maladies endémiques transmises par les moustiques (Ludwig et coll., 2019). Des cas humains de VSGC ont été détectés partout au Canada, et un seul cas de VEEE chez l’humain a été signalé en 2016 en Ontario (M. Drebot, communication personnelle, 2019).
La sensibilisation accrue aux VSGC, l’amélioration de la surveillance sur le terrain des hôtes réservoirs et une plus grande capacité de diagnostic chez les humains et les animaux ont possiblement contribué à leur « émergence » comme problème de santé publique pendant la saison des moustiques. Des tests diagnostiques de routine pour les VSGC étaient réalisés à la fin des années 1970 et au cours des années 1980; ils ont cependant été interrompus jusqu’à ce que de nouvelles méthodes d’essai soient introduites en 2005, lorsque des cas chez l’humain ont encore une fois été documentés au Canada. De 2005 à 2014, plus de 200 cas probables et confirmés d’infections et d’expositions au VSGC ont été dépistés par une surveillance en laboratoire, la maladie causée par le VJC étant détectée plus souvent que celle causée par le VSSH (Drebot, 2015; Lau et coll., 2017; Webster et coll., 2017; M. Drebot, communication personnelle, 2019). Bien que la déclaration des infections par le VSGC ne soit pas obligatoire à l’échelle nationale, le nombre d’infections par ce virus est mentionné dans les rapports annuels sur les arbovirus de l’Agence de la santé publique du Canada (gouvernement du Canada, 2019a) et varie de 34 à 122 cas par année. À ce jour, aucun lien direct n’a été observé entre les effets de la variabilité des conditions météorologiques ou des changements climatiques récents et l’incidence de ces virus transmis par les moustiques au Canada, bien qu’un tel lien puisse exister.
Au Canada, des changements ont été observés au chapitre de la répartition géographique et de la densité des moustiques vecteurs. La faune de moustiques du Canada, qui comprend 74 espèces de moustiques de 10 genres différents, a été décrite dans les années 1970 (Wood et coll., 1979). Depuis, six espèces (Ochlerotatus ventrovittis, Oc. japonicus, Culex salinarius, Cx. erraticus, Anopheles perplexens et An.crucians) ont été déclarées comme étant peut-être nouvellement établies au Canada (Thielman et Hunter, 2007; Giordano et coll., 2015; Iranpour et coll., 2017). De plus, l’aire de répartition géographique de dix espèces (Uranotaenia sapphirina, Culiseta melanura, Cs. minnesotae, Culex tarsalis, Ochlerotatus sticticus, Oc. spencerii, Oc. dorsalis, Oc. nigromaculis, Oc. campestris et Oc. cataphylla) a pris de l’expansion au Canada (Iranpour et coll., 2009). Il est possible que certaines de ces expansions de l’aire de répartition, lesquelles peuvent avoir une incidence sur la santé publique, aient été facilitées par les changements climatiques; toutefois, le manque de surveillance systématique ne permet pas de conclure à un tel lien.
Il y a de solides données d’observation qui étayent des changements au chapitre de l’aire de répartition des espèces de mammifères et d’oiseaux en Amérique du Nord. Au cours des 40 dernières années, environ 180 des 305 espèces d’oiseaux hivernant en Amérique du Nord ont étendu leur aire de répartition vers le nord à un rythme moyen de 1,4 km par année. De même, les aires de reproduction des oiseaux dans le sud de l’Amérique du Nord se sont déplacées de 2,4 km par année en moyenne (gouvernements fédéral, provinciaux et territoriaux du Canada, 2010). Dans les forêts du nord-est de l’Amérique du Nord, 27 des 38 espèces dont l’aire de répartition historique est documentée ont étendu leur aire de répartition, principalement vers le nord (Rodenhouse et coll., 2009). Une évolution de l’aire de répartition au Canada a été recensée pour un certain nombre d’espèces (Hitch et Leberg, 2007; Blancher et coll., 2008;), et des analyses détaillées ont été réalisées pour certaines espèces, notamment la paruline à capuchon (Setophaga citrina) (Melles et coll., 2011) et le petit polatouche (Glaucomys volans) (Garroway et coll., 2010; Garroway et coll., 2011). Il est très possible que les déplacements géographiques de ces espèces soient en partie attribuables au réchauffement climatique récent. De tels changements dans l’aire de répartition pourraient avoir une incidence sur la transmission par les moustiques des agents pathogènes endémiques en modifiant l’aire de répartition des espèces réservoirs, tandis que les espèces qui ne sont pas des réservoirs pourraient « diluer » les cycles de transmission des arbovirus (Levine et coll., 2017). Cependant, d’autres études sont nécessaires pour comprendre précisément comment, où et quand cela peut avoir un impact sur les risques pour les humains.
6.3.1.4 Autres zoonoses transmises par les insectes
Les changements climatiques peuvent également avoir un impact sur d’autres zoonoses transmises par les insectes. Des cas de peste, causée par la bactérie Yersinia pestis, ont été documentés sporadiquement dans l’ouest du Canada. Cette bactérie se transmet par la morsure d’une puce infectée ou par contact direct avec des tissus ou des liquides infectieux lors de la manipulation d’un animal ou d’un être humain qui a contracté la maladie ou qui est décédé des suites de celle-ci. La peste pneumonique peut également être transmise par les gouttelettes produites par la toux ou les éternuements d’un animal ou d’un humain infecté (Centers for Disease Control and Protection, 2019a). Les personnes infectées par la peste développent habituellement des symptômes assimilables à ceux de la grippe. Après cette phase grippale, ils développent des symptômes variables selon la forme de peste – bubonique, septicémique (cette forme se développe généralement comme une complication de la peste bubonique) ou pneumonique. La peste est une infection qui nécessite des soins médicaux d’urgence, car les taux de mortalité sont élevés en l’absence de traitement (Centers for Disease Control and Protection, 2019a). Les réservoirs naturels de Y. pestis sont des rongeurs sauvages, en particulier des spermophiles en Amérique du Nord, et des gerbilles en Asie.
Il y a un lien bien connu entre les régimes climatiques (généralement les périodes plus chaudes et plus humides) et le passage interespèce de la peste des cycles de transmission gerbilles-puces aux humains en Asie centrale en raison des effets sur la végétation qui favorisent les populations de gerbilles et, ensuite, les populations de puces (Kausrud et coll., 2010; Samia et coll., 2011). On pense que ces événements de passage interespèce en Asie ont été la source des grandes pandémies de peste (peste de Justinien, Peste noire) qui ont décimé les populations humaines en Europe (Kausrud et coll., 2010). L’impact du climat a été étudié sur les épidémies de peste dans l’Europe préindustrielle (1347-1760 après J.-C). Contrairement au passage interespèce dans les steppes asiatiques, les résultats semblent indiquer que la peste en Europe était associée à des climats plus secs et plus froids (Yue et Lee, 2018). Cette différence est probablement imputable au fait que la transmission en Europe est due à une combinaison de transmission directe de personne à personne (provoquant une peste de type pneumonique chez l’humain) et de transmission par les puces à partir de réservoirs de rats péridomestiques (produisant la peste de type bubonique), les deux pouvant être influencés par les effets du climat sur la densité et le comportement de la population humaine (Earn et coll., 2020). Une série chronologique de 56 ans de cas de peste chez l’humain dans l’ouest des États-Unis a été utilisée pour étudier les effets des modèles climatiques sur l’incidence de la peste. Comme en Asie centrale, un climat plus chaud et plus humide a été associé à une augmentation du nombre de cas chez l’humain (Ben Ari et coll., 2008). Dans une étude consécutive, le même groupe a trouvé que la combinaison du phénomène El Niño-oscillation australe et de l’oscillation décennale du Pacifique, en favorisant la transition vers un climat plus humide, avait une incidence sur la dynamique de la peste dans l’ouest des États-Unis. Le mécanisme sous-jacent pourrait comprendre des changements dans les précipitations et les températures qui ont une incidence sur les hôtes et les vecteurs comme en Asie. La neige peut jouer un rôle clé, possiblement par des effets sur l’humidité du sol en été, qui a une incidence sur la survie et le développement des puces, et la croissance de la végétation nécessaire aux rongeurs (Ben Ari et coll., 2010). Les résultats d’une étude sur le lien entre les variables climatiques et la fréquence des cas de peste humaine (de 1960 à 1997) dans le nord-est de l’Arizona et le nord-ouest du Nouveau-Mexique portent à croire qu’il est possible d’estimer le risque de peste en surveillant les variables climatiques clés, plus particulièrement les valeurs des températures maximales quotidiennes en été et les quantités de précipitations décalées dans le temps (un et deux ans) en fin d’hiver (février-mars) (Enscore et coll., 2002).
Des études de modélisation ont été menées sur l’impact des changements climatiques sur la distribution de la peste en Amérique du Nord. Basés sur la modélisation des niches écologiques, les modèles de Holt et coll. (2009) prévoient que, d’ici 2050, les conditions climatiques pourraient réduire le risque de peste dans les zones sud de la Californie et accroître le risque le long de la côte nord et dans les Sierras. Une étude réalisée par Nakazawa et coll. (2007) a révélé que la maladie se déplace en fonction des changements climatiques, mais que les changements géographiques globaux seront probablement subtils, avec un certain déplacement des limites sud vers le nord et peut-être un déplacement des limites nord vers le nord également. Des études sur les espèces de puces vectrices Y. pestis dans les populations de chiens de prairie au Canada ont suggéré que le nombre de puces par individu variait inversement au nombre de jours de la saison de croissance précédente ayant reçu plus de 10 mm de précipitations, un indice du nombre d’événements de précipitations qui pourraient avoir causé une augmentation substantielle et prolongée de l’humidité du sol et de la croissance végétative (Eads et Hoogland, 2017). En dehors de ces études, aucune tentative n’a été faite pour évaluer précisément comment les changements climatiques peuvent avoir un impact sur la dynamique de la peste et son aire de répartition géographique au Canada, et aucune donnée de surveillance sur le terrain n’est disponible pour étudier les impacts climatiques sur les aléas environnementaux liés à la peste au Canada. La peste est une maladie à déclaration obligatoire à l’échelle nationale, mais un seul cas chez l’humain, qui a survenu en 1939, a été recensé (gouvernement du Canada, 2018a).
La maladie de Chagas est causée par le parasite protozoaire Trypanosoma cruzi et est une infection qui est le plus souvent contractée par contact avec les matières fécales du triatome (punaise américaine), un insecte hématophage qui se nourrit de sang humain et animal. La maladie de Chagas a une phase aiguë et chronique et, si elle n’est pas traitée, l’infection dure toute la vie. L’infection peut être légère ou asymptomatique. Il peut y avoir de la fièvre et de l’enflure autour du site d’inoculation. Bon nombre de personnes peuvent demeurer asymptomatiques toute leur vie, mais de 20 % à 30 % des personnes infectées développeront tôt ou tard des problèmes médicaux débilitants et parfois mortels (Centers for Disease Control and Prevention, 2018e). La maladie de Chagas n’est pas présente au Canada, et si la plupart des 300 000 cas de maladie de Chagas estimés chez les personnes vivant aux États-Unis ont probablement été acquis dans des pays d’Amérique latine, des cycles de transmission de T. cruzi impliquant des hôtes animaux et des humains, et des infections humaines autochtones transmises par un vecteur ont été signalés dans les États américains du Texas, de la Californie, du Tennessee, de la Louisiane et du Mississippi (Steverding, 2014).
La prévalence de la maladie de Chagas est probablement sous-estimée et son impact se fait sentir sur le système de soins de santé et l’économie mondiale en raison du dépistage et du traitement limités et d’un manque de sensibilisation chez les professionnels de la santé (Click Lambert et coll., 2008; Bern et Montgomery, 2009). L’impact potentiel des changements climatiques sur les vecteurs de la maladie de Chagas en Amérique du Nord a été étudié à l’aide de la modélisation des niches écologiques; bien que l’on ait prédit l’expansion de l’aire de répartition de certaines espèces vers le nord, aucune augmentation des risques n’a été relevée pour le Canada (Carmona-Castro et coll., 2018).
6.3.1.5 Zoonoses transmises par les tiques
Les tiques transmettent une vaste gamme d’agents pathogènes bactériens, viraux et protozoaires à l’échelle mondiale (Sonenshine, 2018). Bien qu’il soit généralement admis que les augmentations de température liées aux changements climatiques contribueront probablement à un accroissement général du nombre, du type, du niveau d’activité et de la répartition géographique des tiques en Amérique du Nord (Eisen et coll., 2016; Sonenshine, 2018), l’ampleur des impacts des changements climatiques sur les risques associés aux maladies transmises par les tiques est incertaine et variera probablement d’une région à l’autre. Au Canada, les données probantes indiquent que l’émergence de la maladie de Lyme associée à la propagation de la tique Ixodes scapularis vers le nord est imputable, du moins en partie, au récent réchauffement climatique (Ebi et coll., 2017; Hoegh-Guldberg et coll., 2019).
Les maladies transmises par les tiques qui ont une incidence sur la santé publique sont des zoonoses et, en Amérique du Nord, les hôtes réservoirs naturels sont des animaux sauvages, en particulier des rongeurs. Il existe deux types de tiques : les tiques dures (Ixodidae) et les tiques molles (Argasidae) (Lindquist et coll., 2016). Dans le nord de l’Amérique du Nord, y compris au Canada, la tique molle la plus importante pour la santé publique est la tique Ornithodorus hermsi, qui transmet la bactérie responsable de la fièvre récurrente, Borrelia hermsii. D’autres tiques molles et espèces Borrelia responsables de la fièvre récurrente sont présentes aux États-Unis; Borrelia turicatae et Borrelia parkeri sont transmises par Ornithodoros turicata et Ornithodoros parkeri, respectivement, mais ne sont pas endémiques au Canada en ce moment (Sage et coll., 2017). Les espèces O. hermsi et B. hermsii vivent naturellement dans des cavernes, et les rongeurs sauvages sont les hôtes naturels dont les tiques se nourrissent et les hôtes réservoirs de B. hermsii. Même si les tiques Ornithodorus hermsi et B. hermsii se trouvent le plus souvent dans les grottes des régions montagneuses de l’ouest des États-Unis et ont une répartition limitée dans le sud de la Colombie-Britannique, ils envahissent les chalets dans ces régions, là où la plupart des cas chez l’humain sont contractés (Dworkin et coll., 2008).
La fièvre récurrente transmise par les tiques est une maladie septicémique fébrile dont l’apparition soudaine est suivie de nombreuses rechutes avec des intervalles afébriles (Artsob, 2000; Murray, 2003). La persistance des bactéries et les rechutes s’expliquent par le fait que la bactérie échappe à la réponse immunitaire (Cutler, 2010). Les symptômes sont très variés : éruptions cutanées, lésions oculaires, jaunisse et vomissements (Ogden et coll., 2014b). Toutefois, elle est rare chez les humains au Canada, en raison de la faible fréquence avec laquelle les gens entrent en contact avec des tiques infectées.
La modélisation de niche écologique a permis de déterminer que l’élévation (les plus élevées étant plus favorables) et des plages précises de température et de précipitations étaient des déterminants clés de la présence des tiques O. hermsi et B. hermsii (Sage et coll., 2017). Cette même étude a projeté l’expansion de l’aire de répartition vers le nord et l’ouest dans les régions montagneuses de la Colombie-Britannique à l’aide de trois modèles climatiques mondiaux (MCM) (ACCESS1-0, HadGEM2-ES et CCSM4) et de deux estimations des trajectoires de concentration de gaz à effet de serre indiquées par les profils représentatifs d’évolution de concentration (RCP) 4.5 et 8.5. La plus grande expansion de l’aire de répartition au Canada était observée avec le scénario d’émissions de gaz à effet de serre plus élevées, RCP 8.5. Tous les modèles basés sur les scénarios RCP 4.5 et 8.5 prévoyaient une contraction de la répartition aux États-Unis.
Les maladies transmises par les tiques dures posent les plus grands défis en matière de santé publique en Amérique du Nord, et parmi ceux-ci, la plus importante est la maladie de Lyme causée par la bactérie Borrelia burgdorferi. La maladie de Lyme est une maladie qui touche plusieurs systèmes corporels et qui commence par une maladie bénigne aspécifique et, dans la plupart des cas, une éruption cutanée typique appelée érythème migrateur. Si elle n’est pas traitée, la maladie progresse à la maladie de Lyme avec des manifestations neurologiques ou cardiaques et, aux stades avancés, à l’arthrite (Wormser et coll., 2006). Cette bactérie est transmise par la tique Ixodes scapularis (la tique à pattes noires) dans le nord-est et le Haut-Midwest des États-Unis, et dans le sud du centre et de l’est du Canada, et par la tique I. pacificus (la tique à pattes noires occidentale) dans les états américains qui bordent le Pacifique et le sud de la Colombie-Britannique (Bouchard et coll., 2015; Eisen et coll., 2016). Dans le sud de la Colombie-Britannique, l’aire de répartition géographique de I. pacificus, et le risque d’infection par B. burgdorferi que pose la tique, serait assez vaste (Mak et coll., 2010). Toutefois, le risque de contracter la maladie de Lyme est beaucoup plus faible là où I. pacificus est le vecteur, par rapport à la plupart des régions où I. scapularis est le vecteur, en raison des caractéristiques de l’écologie de I. pacificus qui font que la prévalence de l’infection chez cette tique est généralement faible et qu’il est moins probable qu’elle pique les humains (Eisen et coll., 2016). Jusqu’à maintenant, aucune étude n’a été menée pour évaluer les impacts possibles des changements climatiques sur les répartitions futures de I. pacificus et de B. burgdorferi en Colombie-Britannique. La modélisation des distributions actuelles de I. pacificus et de B. burgdorferi en Colombie-Britannique a déterminé que les températures moyennes quotidiennes en janvier et en juillet étaient un déterminant important de la niche écologique de ces espèces, et on s’attend à un impact des changements climatiques sur les distributions vers le nord et possiblement en altitude (Mak et coll., 2010).
Concernant la tique I. scapularis, une série d’études sur le terrain et en laboratoire (Ogden, 2014) ont indiqué que le principal impact de la température sur cette espèce touche les taux de développement et l’activité. Les habitats forestiers au Canada fournissent aux tiques des refuges dans lesquels elles sont protégées contre les effets directs des très basses températures hivernales qui autrement les tueraient. Les évaluations fondées sur un modèle du risque d’occurrence de I. scapularis ont utilisé les effets de la température sur les taux de développement, et donc sur la durée du cycle de vie, pour déterminer les seuils de basse température permettant la persistance des populations de tiques autosuffisantes (Ogden et coll., 2005). Ces limites ont maintenant été largement validées par des études de terrain qui, de concert avec l’analyse des données de surveillance passive des tiques, ont établi un schéma spatiotemporel de l’aire de répartition sud-nord qui s’étend des États-Unis au Canada (et maintenant à l’intérieur du Canada), ce qui cadre avec l’hypothèse selon laquelle le récent réchauffement climatique a été un facteur clé (Ogden et coll., 2010; Leighton et coll., 2012; Clow et coll., 2017; Ebi et coll., 2017).
L’augmentation rapide de l’incidence de la maladie de Lyme au Canada, déterminée par la surveillance nationale (Gasmi et coll., 2017), cadre avec l’expansion observée de l’aire de répartition des tiques I. scapularis et avec l’accroissement de la prévalence de l’infection chez les populations de tiques récemment établies (Ogden et coll., 2013; Clow et coll., 2017) (voir la figure 6.5). En raison du degré élevé de concordance entre les études et des données probantes à l’appui des impacts des changements climatiques, il est possible d’affirmer avec une très grande certitude que l’émergence de la maladie de Lyme dans l’est et le centre du Canada est associée au récent réchauffement climatique. L’émergence observée de la maladie de Lyme au Canada correspond aux effets des changements climatiques sur la tique qui en est le vecteur. Cependant, il y a aussi des données probantes selon lesquelles le réchauffement du climat peut influer sur le risque de maladie de Lyme en raison des effets sur d’autres parties du cycle de transmission, en particulier l’abondance et l’aire de répartition géographique des principaux hôtes réservoir, les rongeurs (Simon et coll., 2014).
Évolution du risque de maladie de Lyme au Canada et cas chez l’humain.
Figure 6.5
L’évolution des cas de maladie de Lyme chez l’humain est présentée dans le diagramme à barres. Il est à noter que la maladie de Lyme est devenue une maladie à déclaration obligatoire à l’échelle nationale en décembre 2009. Les données de 2009 et des années précédentes sont fondées sur la présentation volontaire de renseignements provenant des provinces et des territoires où la maladie de Lyme était à déclaration obligatoire.
Source
Gasmi et coll., 2018.
D’autres agents pathogènes transmis par les tiques Ixodes scapularis, en plus de B. burgdorferi, émergent au Canada en provenance des États-Unis à mesure que les tiques se propagent vers le nord (tableau 6.3), mais aucune des maladies dont ils sont responsables n’est actuellement à déclaration obligatoire à l’échelle nationale. Des renseignements détaillés à l’égard de ceux-ci sont présentés dans l’encadré 6.3.
Une évaluation basée sur un modèle des effets potentiels des changements climatiques sur l’aire de répartition géographique de la tique Dermacentor variabilis, un vecteur de la fièvre pourprée des montagnes Rocheuses (FPMR) et de la tularémie, qui est déjà endémique dans une grande partie du sud du Canada, semblait indiquer qu’il y aura une expansion de l’aire de répartition de cette tique vers le nord au Canada (Minigan et coll., 2018). Des températures extrêmes prolongées (c.-à-d. élevées ou basses), une faible humidité et des précipitations intenses pourraient nuire à la survie des tiques en réduisant leur activité et en accroissant leur taux de mortalité (Ogden et Lindsay, 2016). Cependant, on s’attend à ce que cela ait moins d’effet sur les tiques que sur les moustiques en raison de la capacité des tiques à trouver refuge dans leurs habitats boisés (Ogden et Lindsay, 2016). Des études de modélisation semblaient indiquer que, bien que l’on prédise que la limite nord de l’aire de répartition géographique des tiques I. scapularis et D. variabilis se déplacera vers le nord en raison des changements climatiques, il y aura également une contraction vers le nord de la limite sud de l’aire de répartition des tiques à mesure que le climat devient trop chaud pour elles (Brownstein et coll., 2005a; Minigan et coll., 2018). Si c’est le cas, cela peut avoir une incidence sur l’exposition aux tiques pour les personnes vivant dans les États du sud des États-Unis, mais il est peu probable que cela ait une incidence sur le risque au Canada. Des études sur la tique I. scapularis semblent indiquer que les tiques du sud-est des États-Unis pourraient s’adapter à un climat chaud (Ogden et coll., 2018), de sorte que cet effet pourrait être minime.
Les facteurs non climatiques déterminent également où les populations de tiques et les cycles de transmission des agents pathogènes peuvent s’établir. Il s’agit notamment des caractéristiques du microhabitat, comme les caractéristiques du sol, qui sont essentielles à la survie des tiques et à l’établissement réussi de nouvelles populations de tiques (Lindsay et coll., 1998; Guerra et coll., 2002). Les modifications des caractéristiques de l’habitat, parallèlement aux changements climatiques, comme la fragmentation de l’habitat, la perte de biodiversité, la disponibilité des ressources et l’aménagement du territoire, ont une incidence sur la dynamique des tiques, leurs hôtes animaux et l’exposition des humains aux tiques (Brownstein et coll., 2005b; Simon et coll., 2014). Dans certaines régions du Canada, l’émergence des tiques I. scapularis et B. burgdorferi est irrégulière et inégale, probablement en raison du caractère favorable de différents boisés pour la survie des tiques, et en raison des variations pour ce qui est de l’abondance des hôtes des tiques (Gabriele-Rivet et coll., 2015). Les changements au chapitre des facteurs non climatiques peuvent entraîner l’émergence de maladies transmises par les tiques. Par exemple, la maladie de Lyme est apparue aux États-Unis au XXe siècle, probablement en raison du reboisement des terres agricoles et de l’augmentation des populations de cerfs, ce qui a rendu possible l’expansion des populations de tiques (Kilpatrick et coll., 2017).
Bien que la maladie de Lyme et d’autres maladies transmises par les tiques I. scapularis et I. pacificus soient les maladies transmises par les tiques les plus préoccupantes pour la santé publique au Canada à l’heure actuelle, d’autres maladies endémiques transmises par les tiques au Canada (tableau 6.3) pourraient être touchées par le réchauffement du climat ou sont susceptibles de l’être à l’avenir. Il n’y a pas encore de données probantes sur l’impact du réchauffement récent sur ces maladies; toutefois, des données de surveillance indiquent des augmentations récentes de l’abondance et de l’étendue des tiques vectrices de ces autres maladies. Il s’agit notamment de l’abondance accrue de la tique I. cookei (un vecteur de la lignée II du virus Powassan), de la Dermacentor variabilis (un vecteur de la FPMR, de la tularémie et de la fièvre à tiques du Colorado) et de la Rhipicephalus sanguineus (un vecteur de la FPMR) dans l’est du Canada (Gasmi et coll., 2018), et de l’expansion de l’aire de répartition de la tique D. variabilis dans les provinces des Prairies (Dergousoff et coll., 2013) (tableau 6.3).
L’expansion vers le nord de l’aire de répartition de la maladie de Lyme au Canada valide le concept selon lequel :
l’aire de répartition géographique des tiques et des maladies transmises par les tiques en Amérique du Nord peut être limitée par le climat;
il est possible que le réchauffement climatique permette l’expansion de l’aire de répartition de ces espèces vers le nord;
les oiseaux migrateurs et, possiblement, d’autres espèces de mammifères hôtes agissent comme une voie d’introduction de tiques et d’agents pathogènes transmis par les tiques chaque année des États-Unis vers certaines régions du Canada qui deviennent adaptées sur le plan climatique à leur caractère endémique (Ogden et coll., 2008; Nelder et coll., 2019).
Par conséquent, d’autres tiques endémiques aux États-Unis et des agents pathogènes transmis par les tiques qui ne sont pas actuellement endémiques au Canada pourraient se propager vers le nord. L’expansion vers le nord de l’aire de répartition de la tique Amblyomma americanum, un vecteur de la tularémie, du virus Heartland et des agents pathogènes bactériens Ehrlichia ewingii et E. chaffeensis, de même que la cause présumée des allergies à la viande rouge (Reynolds et Elston, 2017), a été observée aux États-Unis (Stafford et coll., 2018). De nombreuses évaluations fondées sur des modèles indiquent que le climat des régions du sud du Canada deviendra adapté pour cette tique en raison du réchauffement climatique (Raghavan et coll., 2019; Sagurova et coll., 2019). Cette tique émergente, de même que les maladies infectieuses et autres problèmes de santé publique qu’elle cause, sont susceptibles de se propager au Canada au cours des prochaines décennies, et seront probablement introduites par des oiseaux migrateurs (Gasmi et coll., 2018; Nelder et coll., 2019).
Tableau 6.3
Agents pathogènes transmis par les tiques qui touchent les humains et espèces de tiques vectrices connexes qui sont présentes ou qui pourraient se propager au Canada
Divers mammifères sauvages, y compris les rongeurs
C.-B., Alb., Sask., Ont. et N.-É.
États de l’est, du centre, de l’ouest et du sud-ouest
Non
Ouib
Oui
Oui
Remarque : * Lorsqu’il n’y a pas de nom de maladie précis, les maladies portent le nom de l’agent pathogène (p. ex., maladie du virus Powassan).
Le symbole « – » indique qu’aucune donnée n’est disponible ou qu’aucune étude n’a été réalisée.
a Canada : Les provinces où la transmission endémique est un fait connu sont soulignées. Quant à celles qui ne le sont pas, les cycles de transmission locaux d’agents pathogènes ne sont pas connus, et des infections ont été détectées chez des tiques adventices, des humains ou des animaux probablement infectés par des tiques adventices.
É.-U. : États où l’incidence de cas chez les humains était la plus élevée.
b Basé sur des enquêtes menées par le passé sur les tiques au Canada, plutôt que sur des enquêtes récentes.
cFrancisella tularensis peut être transmise mécaniquement par une gamme de mouches piqueuses, mais seules les tiques agissent comme vecteurs biologiques.
Zoonoses transmises directement des animaux aux humains
Partager
La présente section porte sur les zoonoses qui peuvent être transmises directement des animaux aux humains (figure 6.6). Les zoonoses transmises dans les aliments et l’eau sont examinées au chapitre 7 : Qualité, quantité et sécurité de l’eau, et au chapitre 8 : Salubrité et sécurité des aliments. Le risque d’infection par les zoonoses transmises directement peut dépendre des effets directs des changements climatiques et des conditions météorologiques sur la survie des agents pathogènes, des effets indirects sur les espèces et les populations hôtes et des effets sur les taux de contact entre les agents pathogènes et les humains. Ensemble, ces facteurs déterminent la fréquence des événements de « passage interespèce », c’est-à-dire lorsque les agents pathogènes sont transmis avec succès des animaux aux humains (Altizer et coll., 2013; Brierley et coll., 2016), de même que la possibilité d’émergence adaptative de zoonoses, qui entraînent ensuite des épidémies ou des pandémies touchant les populations humaines (Ogden et coll., 2017). Les changements climatiques peuvent donc engendrer l’émergence de maladies infectieuses zoonotiques à l’échelle mondiale. On pense que les zoonoses transmises directement, principalement par les espèces sauvages, sont celles qui émergent et réapparaissent le plus souvent à l’échelle mondiale (Jones et coll., 2008).
Les impacts des changements climatiques sur les zoonoses devraient être plus importants dans le Nord qu’ailleurs au Canada, y compris les effets sur les zoonoses d’origine vectorielle, alimentaire et hydrique, de même que les zoonoses transmises directement (Parkinson et coll., 2014). Un impact climatique plus important, conjugué, dans de nombreux cas, à une consommation plus élevée d’aliments traditionnels et prélevés dans la nature1, donne à penser que les risques d’effets sur la santé seront plus importants dans les collectivités nordiques, en particulier les collectivités autochtones du Nord (Brook et coll., 2009). Un autre problème est lié au fait que, à mesure que fond le pergélisol en raison d’un climat plus chaud, des agents pathogènes qui sont demeurés dormants, mais viables dans les carcasses d’animaux ou le sol, peuvent être libérés dans l’environnement et causer des éclosions de maladies (Revich et coll., 2012). Pour la plupart des zoonoses transmises directement dont il est question dans la présente section, des données étayant la sensibilité aux facteurs climatiques et aux conditions météorologiques soulèvent la possibilité d’impacts actuels et futurs des changements climatiques. Dans de très rares cas, on a tenté d’évaluer de tels impacts. En outre, très peu de zoonoses transmises directement présentent des signes de changement au chapitre des tendances en matière de maladies ou de risques, mais cela peut être imputable, en partie, au manque de surveillance systématique de ces maladies au Canada.
Zoonoses et émergence par changement génétique et/ou évolution.
6.3.2.1 Zoonoses virales
À l’échelle mondiale, les virus de la grippe aviaire A représentent l’un des groupes de zoonoses virales transmises directement les plus hautement pathogènes, et leurs impacts sont considérables. Des épidémies mondiales de virus de la grippe aviaire hautement pathogènes (pour les humains) sont survenues au cours des dernières décennies (Goneau et coll., 2018), et des éclosions se sont produites dans la volaille domestiquée au Canada, ce qui a fait craindre l’apparition de cas chez l’humain (Skowronski et coll., 2007). Chez les humains, la maladie varie d’une maladie légère à une maladie grave, avec des taux élevés de mortalité (Neumann, 2015). Les principaux réservoirs des virus de la grippe aviaire sont les oiseaux sauvages, en particulier le gibier d’eau (c.-à-d. les cygnes, les oies et les canards). Les virus de la grippe porcine sont également très préoccupants sur le plan de la santé publique, bien que ces virus soient maintenant principalement établis chez le porc domestiqué, et que les impacts des changements climatiques soient relativement peu probables (Schultz-Cherry et coll., 2011) Les virus pandémiques de la grippe, comme les virus H1N1 de 1918 et de 2009, émergent à la suite de la recombinaison (un processus moléculaire) des virus de la grippe aviaire, porcine et humaine (Neumann et coll., 2009). En raison du niveau élevé de menace que représentent les virus zoonotiques et pandémiques de la grippe, il y a une surveillance approfondie de ces virus chez les animaux sauvages et domestiqués et les humains à l’échelle mondiale, de même qu’au Canada (gouvernement du Canada, 2017; gouvernement du Canada, 2018b), et un programme de préparation aux pandémies est en place (gouvernement du Canada, 2019b).
L’émergence et la réémergence des risques de grippe aviaire chez les humains pourraient être imputables à un certain nombre d’effets des changements climatiques sur l’écologie des virus, y compris les habitudes de migration des oiseaux et les changements dans l’aménagement du territoire et les modes de production du bétail (Morin et coll., 2018). La transmission des virus de la grippe aviaire chez les oiseaux sauvages, qui est souvent une transmission fécale-orale par l’eau, peut être inhibée par le réchauffement climatique, car la survie des virus est meilleure dans l’eau froide que dans l’eau chaude (Morin et coll., 2018). Dans le même ordre d’idées, la prévalence des oiseaux infectés était plus élevée, selon une étude canadienne, si l’échantillonnage était précédé d’une période de temps froid (Papp et coll., 2017), et les données phylogénétiques semblent indiquer que les virus de la grippe A ont en fait émergé après le petit âge glaciaire (Gatherer, 2010). Toutefois, la complexité associée à la transmission mondiale des virus de l’influenza aviaire signifie que, même si les virus de l’influenza aviaire sont intrinsèquement sensibles au climat, les changements climatiques pourraient avoir des répercussions positives en réduisant la transmission de la maladie en raison des effets sur la survie des virus dans l’environnement, ou négatives en accroissant la transmission par les effets indirects d’autres facteurs (Gilbert et coll., 2008; Gatherer, 2010; Morin et coll., 2018).
La rage est une maladie d’origine zoonotique qui représente un problème de santé mondial majeur, avec plus de 55 000 décès chaque année (Knobel et coll., 2005). Le virus de la rage est capable d’infecter les tissus du cerveau et du système nerveux de tous les mammifères, ce qui entraîne habituellement des modifications des comportements suivies de la mort (OMS, 2013). Au Canada, la rage est associée à plusieurs espèces sauvages, dont les chauves-souris, les mouffettes, les ratons laveurs et les renards, qui maintiennent des variants viraux distincts circulant de façon endémique au sein de leurs populations (Rosatte, 1988; Tinline et Gregory, 2020). Les cas humains de rage sont rares au Canada, avec seulement trois cas contractés au pays depuis 2000 (Filejski, 2016), mais l’exposition humaine aux animaux enragés suivie de l’administration d’un traitement prophylactique post-exposition se produit chaque année dans tout le pays.
Bien que les changements écologiques causés par les facteurs climatiques soient susceptibles d’avoir une incidence sur la circulation de la rage dans chacune de ses populations hôtes respectives dans une certaine mesure, ces impacts seront probablement ressentis d’abord et avant tout dans l’Arctique. La rage sévit dans tout l’Arctique et est considérée comme enzootique – ou prévalente – dans le nord du Canada, c’est-à-dire au nord du 60e parallèle (voir le chapitre 2 : Changements climatiques et santé des Autochtones du Canada). Le principal hôte réservoir de la rage dans l’Arctique est le renard arctique (Vulpes lagopus).
La rage dans l’Arctique est causée par une souche unique du virus de la rage appelée « variant arctique du virus de la rage », qui circule dans les régions arctiques et subarctiques avec des incursions sporadiques vers les régions plus au sud, ce qui mène parfois à l’établissement de la rage enzootique dans ces régions (Rosatte, 1988). Dans l’Arctique canadien, les cas de rage chez les animaux domestiques et les animaux sauvages sont signalés chaque année, et les résidents de l’Arctique reçoivent plus de traitements prophylactiques contre la rage par habitant que toute autre population au Canada (Rosatte, 1988; Mitchell et Kandola, 2005; Aenishaenslin et coll., 2014; ACIA, 2019).
Les changements climatiques sont plus rapides et plus vastes dans le nord du Canada que les moyennes mondiales et canadiennes (Larsen et coll., 2014; Bush et Lemmen, 2019). Bien que les impacts à long terme du réchauffement rapide du climat sur la rage dans l’Arctique n’aient pas encore été documentés, l’écologie et l’épidémiologie de la rage risquent d’être modifiées par les perturbations continues des écosystèmes arctiques causées par le climat. Il s’agit notamment des cycles de décroissance des populations de lemmings, une source essentielle de nourriture pour les renards arctiques (Fuglei et Ims, 2008; Kausrud et coll., 2008), de la réduction de la durée et de l’étendue de la glace de mer utilisée par les renards pour l’alimentation et les déplacements hivernaux (Kim et coll., 2016), et de l’expansion de l’aire de répartition des renards roux (Vulpes vulpes) vers le nord. Les renards roux font compétition aux renards arctiques pour les ressources et sont un hôte réservoir compétent pour le variant de la rage dans l’Arctique (Rosatte, 1988; Hersteinsson et MacDonald, 1992; Gallant et coll., 2012). Selon une étude de modélisation récente (Simon et coll., 2019), l’incidence de la rage pourrait d’abord se stabiliser en raison de la réduction de la variation de la dynamique des proies, les interactions entre les renards arctiques et roux intensifiant les éclosions là où ces espèces se chevauchent. Toutefois, à long terme, le déplacement des renards arctiques par les renards roux et la baisse d’activité hivernale en raison de la réduction de l’étendue de la glace de mer peuvent limiter la transmission de la rage dans les régions plus chaudes. De même, Huettmann et coll. (2017) ont prédit une réduction de la niche écologique actuelle du virus de la rage en Alaska avec le réchauffement climatique.
Les hantavirus causent des infections dans les populations de rongeurs sauvages et de mammifères insectivores et peuvent entraîner le syndrome pulmonaire à hantavirus, une fièvre suivie d’un œdème pulmonaire aigu et de l’entrée en état de choc. Il n’y a pas de traitement particulier pour le syndrome pulmonaire à hantavirus; le taux de mortalité est de 38 % au Canada (Drebot et coll., 2000), où on signale en moyenne quatre cas chaque année (gouvernement du Canada, 2015). Les humains contractent l’infection par le contact avec des rongeurs infectés ou leurs excréments, en particulier l’urine ou les fèces aérosolisées (Weir, 2005). Dans un rapport précédent (Charron et coll., 2008), il a été déterminé que l’incidence des hantavirus est sensible aux facteurs climatiques et est associée aux effets sur les populations de rongeurs. Il n’y a pas eu d’autres études sur les hantavirus au Canada, et les impacts potentiels des changements climatiques sont inexplorés.
6.3.2.2 Zoonoses parasitaires
Au Canada, les protozoaires zoonotiques Cryptosporidium, Giardia et Toxoplasma sont les parasites les plus importants pour la santé publique, selon l’opinion d’experts, et les deux premiers sont à déclaration obligatoire à l’échelle nationale (ONUAA et OMS, 2014). Les protozoaires Cryptosporidium et Giardia spp. peuvent être transmis directement par les animaux, par exemple, les animaux de ferme nouveau-nés ou les animaux des zoos pour enfants. Il peut également y avoir transmission d’une personne à une autre par l’intermédiaire de contacts étroits et d’éclosions dans des milieux communautaires ou de soins, ou par les voies alimentaire ou hydrique. Les changements climatiques auront une incidence sur les infections par ces parasites en modulant la transmission d’origine alimentaire et hydrique (voir le chapitre 7 : Qualité, quantité et sécurité de l’eau et le chapitre 8 : Salubrité et sécurité des aliments).
À l’heure actuelle, le fardeau humain de la maladie, y compris les lésions neuronales et oculaires associées à la migration larvaire viscérale (c.-à-d. la migration des larves dans le corps) des ascarides zoonotiques (c.-à-d. les vers ronds) Baylisascaris procyonis et Toxocara canis, n’est pas connu au Canada, et seuls les cas isolés et les données d’enquêtes de sérosurveillance sont signalés (Embil et coll., 1988; Sapp et coll., 2016). Les hôtes réservoirs de ces ascarides sont, respectivement, les ratons laveurs et les chiens domestiques. Dans le sud du Canada, les chiens sont aussi des hôtes compétents pour les ascarides B. procyonis (Lee et coll., 2010). Une étude nationale au Canada a rapporté que le parasite Toxocara canis était celui qui était le plus souvent détecté dans les fèces des chiens dans les refuges (Villeneuve et coll., 2015). La transmission directe des chiens aux humains est possible par contact avec des œufs embryonnés présents sur la fourrure de l’animal, mais en général, on pense que le parasite T. canis est principalement acquis à partir d’œufs embryonnés présents dans l’environnement (Keegan et Holland, 2010).
Les œufs de ces deux ascarides passent par un stade de développement dans l’environnement qui est dépendant de la température avant de devenir infectieux pour les humains. Par conséquent, les principaux mécanismes par lesquels les changements climatiques peuvent avoir une incidence sur ces parasites sont l’amplification locale due à un développement plus rapide et à une survie accrue des œufs déposés dans les fèces des hôtes définitifs (c’est-à-dire ceux dans lesquels la reproduction du parasite a lieu), et l’expansion vers le nord de l’aire de répartition en raison des changements dans la distribution et l’abondance des hôtes canins et des ratons laveurs. Les œufs de T. canis sont sensibles au gel (O’Lorcain, 1995) et leur développement est retardé après refroidissement, mais les œufs de B. procyonis peuvent survivre aux températures de congélation et aux cycles de gel-dégel (Shafir et coll., 2011). Cela porte à croire qu’un climat en évolution peut avoir différents effets sur ces parasites. Pour le parasite B. procyonis, l’expansion vers le nord pourrait suivre celle de l’hôte principal, le raton laveur, dont l’aire de répartition est actuellement limitée aux parties du sud des provinces canadiennes (Naughton, 2012). Dans le cas du T. canis, l’expansion de l’aire de répartition vers le nord et l’amplification locale peuvent suivre l’augmentation du taux de survie et de développement des œufs dans l’environnement. L’augmentation de la circulation des deux parasites chez les chiens peut entraîner une augmentation des maladies chez les Canadiens et les Canadiennes en raison de leur lien étroit. Jusqu’à maintenant, aucune étude n’a examiné explicitement les effets des changements climatiques sur ces parasites. Toutefois, des œufs de T. canis ont récemment été signalés au Canada chez des chiens au nord du 60e parallèle pour la première fois (Salb et coll., 2008), ce qui pourrait indiquer une expansion de l’aire de répartition de cette espèce vers le nord (Jenkins et coll., 2011).
Jusqu’à récemment, on pensait que le cestode zoonotique Echinococcus multilocularis n’était transmis entre les canidés sauvages et les rongeurs que dans les régions du sud des provinces des Prairies et de la toundra arctique, et un seul cas humain autochtone avait été documenté au Canada (James et Boyd, 1937; Deplazes et coll., 2017). Les œufs d’Echinococcus sont excrétés dans les fèces des canidés sauvages ou domestiques, sont immédiatement infectieux et peuvent adhérer à la fourrure de l’hôte. L’infection peut se produire chez l’humain lorsqu’il consomme ces œufs par inadvertance. La transmission peut également se produire par la consommation de produits non lavés ou la consommation d’eau de surface non filtrée dans des environnements fortement contaminés (Deplazes et coll., 2017). Chez les humains, le parasite s’établit d’abord dans le foie et se comporte comme une tumeur parasitaire, finissant par produire des métastases dans tout l’abdomen, à moins d’une intervention chirurgicale et médicale agressive. Plus de cas d’échinococcose alvéolaire ont récemment été détectés chez les chiens et les humains au Canada, et le parasite a élargi son aire de répartition pour inclure la majeure partie de l’Ouest canadien (Deplazes et coll., 2017) et la partie sud de l’Ontario (Kotwa et coll., 2019). Cette émergence est probablement liée à la fois à l’introduction de souches européennes du parasite, qui se sont maintenant établies avec succès chez les espèces sauvages, et à l’accroissement des taux de contact entre les humains et la faune urbaine.
Les mécanismes possibles des répercussions des changements climatiques sur ce parasite comprennent des effets sur la survie des œufs, car ceux-ci sont résistants à l’environnement, mais sensibles aux températures élevées et à la dessiccation, de même que la stabilisation et l’amplification des populations d’hôtes intermédiaires (rongeurs), ce qui était considéré comme un facteur limitant le contrôle de ce parasite (Rausch, 1956; Jenkins et coll., 2013). Une corrélation a été établie entre les variables climatiques, à la fois la température et les précipitations, et la prévalence de l’infection chez les hôtes définitifs et intermédiaires (c.-à-d. les hôtes où il n’y a pas de reproduction parasitaire) (Takeuchi-Storm et coll., 2015), qui peut être associée à des effets directs sur la survie des œufs dans l’environnement ou des impacts sur les hôtes réservoirs (Mas-Coma et coll., 2008). Le parasite fait maintenant l’objet d’une déclaration obligatoire provinciale en Alberta et en Ontario. Les chiens peuvent servir d’hôtes définitifs et intermédiaires pour E. multilocularis et, par conséquent, un traitement régulier à l’aide d’un médicament anthelminthique administré aux chiens dont la consommation d’hôtes intermédiaires de rongeurs est connue peut aider à atténuer le risque pour l’humain (Jenkins, 2017).
6.3.2.3 Zoonoses bactériennes
Les bactéries de l’espèce Brucella, qui sont endémiques au Canada et qui causent des maladies chez les humains, sont Brucella abortus et B. suis biotype 4. On retrouve la bactérie Brucella abortus dans le bétail, lorsque la prévention de la maladie n’est pas assurée au moyen de programmes de lutte, et les ongulés sauvages comme le cerf et le bison (Nishi et coll., 2002). La bactérie Brucella suis biotype 4 s’attaque au caribou (Forbes, 1991). Chez les humains, la brucellose cause une fièvre persistante et souvent récurrente accompagnée de fatigue chronique et d’autres symptômes comme l’arthrite (Centers for Disease Control and Prevention, 2019b). Chez les animaux, le principal effet de l’infection est l’avortement. La principale voie de transmission vers la population humaine est d’origine alimentaire, mais les personnes qui travaillent dans les industries du bétail et de la transformation des aliments peuvent la contracter par contact direct avec des tissus du bétail infecté (Hunter et coll., 2015), tout comme les chasseurs peuvent la contracter par contact direct avec des tissus d’animaux sauvages infectés (Franco-Paredes et coll., 2017). Mis à part leurs effets directs possibles sur la survie de la bactérie Brucella spp. dans l’environnement, et donc sur la transmission d’animaux à animaux, les changements climatiques peuvent avoir une incidence sur la transmission par des effets sur la dynamique des populations d’hôtes sauvages (Cross et coll., 2007). Bien que la brucellose puisse être une maladie sensible au climat, aucune étude n’a été réalisée pour explorer les effets possibles des changements climatiques.
Causé par la bactérie Bacillus anthracis, le risque d’anthrax dans l’environnement est amplifié par les ongulés sauvages et domestiqués infectés qui souffrent de maladies hémorragiques à la suite d’une infection. Lorsque l’animal infecté meurt, son sang et sa carcasse contaminent les sols avec des bactéries qui forment des spores résistantes et persistantes (Valseth et coll., 2017). Les humains peuvent contracter l’infection par l’inhalation des spores, par l’ingestion des spores qui contaminent la nourriture ou l’eau, ou par l’introduction des spores dans la peau par le biais d’une coupure.
Les manifestations chez les humains dépendent de la voie de l’infection – manifestations cutanées après l’infection des coupures, manifestations gastro-intestinales après l’ingestion et manifestations respiratoires après l’inhalation. Le taux de décès est élevé (environ 25 %) chez les personnes non traitées (Centers for Disease Control and Prevention, 2017). Des températures élevées ont été associées à des éclosions chez des animaux sauvages et domestiqués dans des pays du Nord, dont le Canada, et on s’attend à ce que l’environnement devienne plus favorable à la transmission de l’anthrax et à la survie de la bactérie B. anthracis dans le nord et le centre-sud du Canada en raison du réchauffement climatique (Walsh et coll., 2018). La nature persistante des spores de B. anthracis dans les sols permet l’apparition d’épidémies causées par les conditions météorologiques qui peuvent devenir plus fréquentes avec les changements climatiques en raison des effets combinés du réchauffement et des inondations (Maksimovic et coll., 2017). L’anthrax et la brucellose sont des maladies à déclaration obligatoire lorsqu’elles sont dépistées dans le bétail au Canada (gouvernement du Canada, 2019c).
La fièvre Q est causée par la bactérie Coxiella burnetii. Cette bactérie est principalement transmise aux humains par voie aérienne dans les élevages de ruminants infectés, en particulier les élevages de moutons et de chèvres. Chez les humains, l’infection cause de la fièvre ainsi qu’une pneumonie, des infections du foie et des infections chroniques du cœur, et le taux de mortalité sans traitement est élevé (jusqu’à 25 %) (Centers for Disease Control and Prevention, 2019c). Chez les ruminants, il y a peu de manifestations cliniques au-delà de l’avortement si l’animal est infecté à la fin de la grossesse. Les animaux infectés excrètent les bactéries dans le lait, l’urine, les fèces, le placenta et les liquides de naissance (Plummer et coll., 2018). La bactérie persiste dans l’environnement; bien que les travailleurs agricoles soient les plus à risque, de grandes éclosions associées à la transmission aérienne sur de longues distances à partir des fermes touchées ont eu lieu en Europe (Schneeberger et coll., 2014). La fièvre Q n’est pas à déclaration obligatoire à l’échelle nationale au Canada, mais on pense qu’elle est particulièrement répandue dans les fermes du Québec (Dolcé et coll., 2003), bien que la raison en soit inconnue. Une incidence accrue a été associée à une augmentation des précipitations dans les Caraïbes, ce qui semble indiquer une sensibilité aux conditions météorologiques (Eldin et coll., 2015; Sivabalan et coll., 2017). La dispersion de la bactérie augmente avec la vitesse du vent et un faible taux d’humidité, mais la réduction des précipitations associées aux changements climatiques dans certaines régions peut réduire la concentration de cet agent pathogène en suspension dans l’air (van Leuken et coll., 2016). Il n’existe aucune étude sur les effets possibles des changements climatiques.
6.3.3
Maladies infectieuses transmises directement d’une personne à une autre
Partager
La présente section est axée sur le sous-ensemble d’infections qui sont transmises directement d’une personne à une autre. La transmission directe peut être définie comme la transmission d’un agent infectieux entre des personnes par contact direct ou par des gouttelettes respiratoires plutôt que par des aliments, l’eau ou des arthropodes vecteurs. Le contact direct se produit par contact peau à peau, tandis que la propagation par les gouttelettes fait référence aux aérosols relativement gros et de courte portée produits par les éternuements et la toux ou en parlant (Dicker et coll., 2012). Les infections transmises directement causent un large éventail de maladies, et de nombreuses caractéristiques de la population humaine – comme l’évolution de la démographie et des comportements humains, les modèles de connectivité et le respect des recommandations en matière de santé publique (p. ex., vaccination contre la grippe) – ont un effet sur le fardeau de ces maladies au sein de la population (Heesterbeek et coll., 2015).
Le taux de reproduction de base (R0) (c.-à-d. le nombre de cas secondaires causés par un cas de maladie primaire au sein d’une population totalement vulnérable et en l’absence d’intervention) est une mesure importante de la transmissibilité (Pandemic Influenza Outbreak Research Modelling Team, 2009). Dans le cas des agents pathogènes transmis directement, le R0 est fonction de la durée de l’infection, du taux de contact effectif d’une personne infectée et de la « contagiosité » d’un cas individuel (Pandemic Influenza Outbreak Research Modelling Team, 2009). Si l’un ou l’autre de ces facteurs est modifié directement ou indirectement par les changements climatiques, il en résultera un changement dans l’épidémiologie des maladies. L’augmentation du R0 peut alourdir le fardeau des maladies endémiques et accroître les risques d’éclosion et d’émergence de maladies. En fait, les impacts des changements climatiques sur l’habitat humain, la productivité agricole, les conflits, les moyens de subsistance et la stabilité politique entraîneront probablement des déplacements de populations et des modifications des schémas de mélange de populations qui auront des répercussions sur les maladies transmissibles; le lien entre les conflits humains, les déplacements de populations, les migrations à grande échelle et les maladies transmissibles a été démontré à maintes reprises dans un passé proche et lointain (Rabaan, 2019). Ces impacts de deuxième et de troisième ordre des changements climatiques sur les maladies transmissibles peuvent éclipser les changements de premier ordre dans l’épidémiologie en raison des impacts sur les hôtes et les agents pathogènes.
De nombreuses études ont cerné les impacts des conditions météorologiques et climatiques sur les maladies infectieuses transmises directement d’une personne à une autre, mais les études qui tentent d’analyser explicitement les effets des changements climatiques font défaut. Seuls les effets pour la maladie mains-pieds-bouche (MMPB) ont été étudiés jusqu’à maintenant, et aucun effort n’a été déployé pour attribuer les changements survenus dans l’incidence de la maladie aux changements climatiques récents.
Un indicateur possible de la modification liée aux facteurs climatiques de l’épidémiologie des maladies transmissibles directement est leur saisonnalité. La saisonnalité de ces infections est bien connue et a fait son chemin dans le langage courant (p. ex., « saison de la grippe »). Si les mécanismes qui sous-tendent le caractère saisonnier des maladies transmissibles restent mal compris (Fisman, 2007; Greer et coll., 2008; Fisman, 2012), un nombre croissant d’ouvrages ont cherché à distinguer la contribution des facteurs environnementaux à la transmission des maladies de celle d’autres facteurs saisonniers, tels que les comportements humains saisonniers (fréquentation scolaire, vacances et rassemblement à l’intérieur) et les facteurs neuro-endocriniens (p. ex., mélatonine, vitamine D) (Dowell, 2001). De nombreux agents pathogènes viraux et bactériens, en particulier ceux qui causent une infection respiratoire et gastro-intestinale, présentent une saisonnalité marquée dans des climats tempérés comme celui du Canada, ce qui indique que les facteurs environnementaux sont d’importants déterminants du risque de maladie, et que les changements environnementaux peuvent entraîner des changements importants dans l’épidémiologie des maladies.
6.3.3.1 Infections respiratoires
Les infections respiratoires sont le facteur qui contribue le plus au fardeau global de la maladie dans le monde (Ferkol et Schraufnagel, 2014); elles représentaient la troisième cause de décès dans le monde en 2015 (Global Burden of Disease, 2015; Mortality and Causes of Death Collaborators, 2016). À cela s’ajoute maintenant la COVID-19; toutefois, à ce jour, il n’y a aucune preuve de l’incidence des conditions météorologiques et climatiques sur l’émergence et la propagation de cette maladie. Les maladies respiratoires transmissibles comprennent les maladies causées par des agents pathogènes viraux et bactériens. Pour de nombreuses maladies respiratoires d’origine virale, comme la grippe, un large éventail de manifestations sont courantes; la plupart des gens souffrent d’une maladie légère à modérée qui guérit sans traitement. Toutefois, une proportion de la population sera plus gravement malade ou développera une infection bactérienne secondaire qui nécessitera une hospitalisation. On estime qu’au Canada, une moyenne de 12 200 hospitalisations (Schanzer et coll., 2006; Schanzer et coll., 2008; Schanzer et coll., 2013a; Schanzer et coll., 2013b) et environ 3 500 décès sont imputables à la grippe chaque année (Schanzer et coll., 2013b). Les infections respiratoires graves sont plus fréquentes aux extrêmes d’âge (jeune et vieux) (Ampofo et coll., 2006; Schanzer et coll., 2006; Schanzer et coll., 2008; Schanzer et coll., 2013a; Schanzer et coll., 2013b).
Dans les régions tempérées, des infections respiratoires virales, comme la grippe et le virus respiratoire syncytial (VRS), peuvent se produire tout au long de l’année, mais on observe un pic épidémique saisonnier pendant les mois d’hiver (Thompson et coll., 2006; Rohr et coll., 2011). Le caractère saisonnier prévisible des infections respiratoires virales hivernales a amené les chercheurs à évaluer si les conditions environnementales pouvaient influer sur le moment de l’apparition ou la gravité des épidémies saisonnières de grippe. Certaines données indiquent que la baisse de l’humidité absolue (Shaman et coll., 2010), la baisse de la température (Earn et coll., 2012; Skog et coll., 2014), ou les deux (Chattopadhyay et coll., 2018) peuvent déclencher le début de la saison de la grippe dans les pays tempérés. La baisse des températures et la diminution de l’humidité absolue peuvent également accroître la transmissibilité de la grippe et des maladies semblables pendant les épidémies (Tang et coll., 2010; Roussel et coll., 2016; Caini et coll., 2018).
La gravité des saisons de la grippe est réduite en présence de fortes conditions du phénomène El Niño-oscillation australe (Viboud et coll., 2004; Choi et coll., 2006), ce qui fournit d’autres données probantes qui étayent la sensibilité au climat. Cette inversion thermique dans l’océan Pacifique est associée à des conditions inhabituellement chaudes et pluvieuses semblables à celles prévues dans les scénarios de changements climatiques (Fisman et coll., 2016). Compte tenu de l’augmentation apparente de la transmission de la grippe par temps froid, on pourrait s’attendre à ce que les changements climatiques aient pour effet de réduire le fardeau de la maladie associée aux infections respiratoires virales. Cependant, les effets du climat sur l’épidémiologie de la grippe peuvent dépendre de l’échelle temporelle. Une analyse du climat et des épidémies de grippe saisonnière antérieures aux États-Unis de 1997 à 2013 a révélé que des hivers doux étaient associés à des épidémies précoces et graves au cours de l’année suivante (Towers et coll., 2013). La diminution de la transmission de la grippe au cours d’une saison donnée peut donner lieu à une hausse du nombre de personnes vulnérables et, par conséquent, à une épidémie de grippe plus explosive au cours de la saison suivante. En outre, il a été suggéré que l’augmentation des températures mondiales pourrait causer un accroissement du taux de diversification de la grippe, ce qui pourrait diminuer l’immunité protectrice et donner lieu à des pandémies de grippe plus fréquentes (Gatherer, 2010).
Plusieurs agents pathogènes respiratoires humains bactériens importants présentent un caractère saisonnier stéréotypé. Les infections à Streptococcus pneumoniae et à Neisseria meningitidis, qui peuvent être causées en partie par un risque accru en raison d’une grippe antérieure et d’une infection par le VRS, sont plus courantes en hiver (Tuite et coll., 2010; Kuster et coll., 2011). Des impacts environnementaux directs sur ces infections sont également possibles. Les infections pneumococciques invasives ont tendance à être fortement saisonnières dans les climats tempérés, ce qui entraîne des corrélations causales avec le froid (Cilloniz et coll., 2017). Cependant, il a été suggéré que la diminution du rayonnement UV hivernal, qui est moins susceptible d’être touché par les changements climatiques que les régimes de température ou de précipitations, pourrait expliquer cette saisonnalité hivernale (White et coll., 2009). Des travaux de modélisation plus récents indiquent que la saisonnalité des infections pneumococciques peut être complexe et déterminée par les recoupements entre les profils de contact de la population, les conditions météorologiques hivernales et les infections respiratoires virales prédisposantes (Domenech de Cellès et coll., 2019). Les infections à méningocoques ont toujours été très saisonnières en Afrique subsaharienne, l’apparition de l’épidémie étant liée à la période des vents de Harmattan (Sultan et coll., 2005). Dans les régions tempérées, l’incidence des infections invasives à méningocoques atteint souvent un sommet à la fin de l’hiver ou au début du printemps (Rosenstein et coll., 2001; Lindsay et coll., 2002; Brachman et Abrutyn, 2009). Cependant, les liens avec les facteurs environnementaux, bien qu’ils soient déterminés dans des régions individuelles, ne sont pas les mêmes d’une région à l’autre (Collier, 1992; Lindsay et coll., 2002; Sultan et coll., 2005; Michele et coll., 2006; Kinlin et coll., 2009). On prévoit que la fréquence accrue des événements météorologiques extrêmes, y compris les chutes de pluie abondantes, accompagnera les changements climatiques, et l’augmentation des maladies respiratoires a été associée à l’exposition aux eaux de crue (De Man et coll., 2016). Toutefois, il est probable que ces maladies soient imputables à des agents pathogènes d’origine environnementale (p. ex., Legionella, tel qu’il est décrit ci-dessous) et à des agents pathogènes d’origine hydrique, qui sont abordés ailleurs dans le présent rapport (voir le chapitre 7 : Qualité, quantité et sécurité de l’eau).
6.3.3.2 Infections gastro-intestinales
Comme pour les infections virales respiratoires, les infections virales du tractus gastro-intestinal sont très saisonnières. Les infections entériques, y compris l’échovirus, les virus coxsackie A et B, et, avant qu’il ne soit maîtrisé, le poliovirus, sont associés à des éclosions à la fin de l’été et au début de l’automne dans les pays tempérés (Moore, 1982). Les norovirus et les rotavirus sont courants en hiver (Greer et coll., 2009; Patel et coll., 2013). Les entérovirus sont habituellement transmis par voie fécale-orale et peuvent causer des maladies reconnaissables, notamment la MMPB, les infections neuroinvasives (p. ex., paralysie flasque aiguë) et des syndromes aspécifiques, y compris la méningite virale, l’encéphalite, la myocardite, la gastroentérite et la conjonctivite. Les norovirus et les rotavirus sont associés à la gastroentérite et les norovirus, en particulier, peuvent causer d’importantes éclosions de sources ponctuelles, y compris celles qui surviennent dans des milieux fermés comme les navires de croisière et les établissements de soins de longue durée (Rajagopalan et Yoshikawa, 2016; Mouchtouri et coll., 2017).
Les mécanismes qui sous-tendent la saisonnalité des infections aux entérovirus ne sont pas entièrement compris. Cependant, Dowell (2001) a souligné que le caractère saisonnier de la poliomyélite devient plus évident avec la distance par rapport à l’équateur, et qu’il ne paraît pas dans les régions périéquatoriales, ce qui laisse entendre que les changements saisonniers des conditions météorologiques sont des facteurs clés de la saisonnalité des entérovirus. Il a été suggéré que la température module la reproduction et la survie des particules virales, et que l’humidité modifie la fixation des particules virales aux gouttelettes d’eau, facilitant ainsi la transmission (Patz et coll., 2005; Wong et coll., 2010). Conformément à ces hypothèses, des températures élevées ont été associées à un risque accru de MMPB dans plusieurs pays d’Asie (e.g., Hii et coll., 2011; Onozuka et Hashizume, 2011), une à deux semaines plus tard. Alors que certaines études semblent montrer que le lien entre la température et le risque de maladie est linéaire (Hii et coll., 2011; Wang et coll., 2016), ou associé à des températures quotidiennes maximales élevées (Sumi et coll., 2017), d’autres semblent révéler des liens non linéaires plus complexes entre le risque et la température (Kim et coll., 2016; Liao et coll., 2016; Zhao et coll., 2018).
L’augmentation de l’humidité relative et des précipitations a également été associée à un risque élevé de MMPB dans la plupart des études (Hii et coll., 2011; Onozuka et Hashizume, 2011; Zhao et coll., 2017), l’humidité pouvant moduler les effets de la température (Kim et coll., 2016; Sumi et coll., 2017). Il a été avancé que les événements de précipitations extrêmes pourraient accroître le risque de MMPB (Cheng et coll., 2014; Yu et coll., 2019). Tel qu’il est susmentionné, la combinaison d’une température plus élevée et de fortes pluies est liée au phénomène El Niño-oscillation australe, et ce phénomène entraîne des augmentations à court terme des cas de MMPB en Chine. Ces observations ont permis de paramétrer des modèles mathématiques qui semblent indiquer une augmentation de 5 % de l’incidence de la MMPB en Chine d’ici 2090, ce qui représenterait un accroissement substantiel du fardeau de la maladie (Zhao et coll., 2018). Si de telles estimations s’appliquent aux infections entérovirales de façon générale, cela pourrait également présenter une augmentation des manifestations actuellement rares, mais graves, de maladies causées par des entérovirus, telles que les infections neuroinvasives et la myocardite.
La hausse des températures et les conditions humides associées aux changements climatiques pourraient donc engendrer une incidence plus forte des maladies entériques endémiques à l’échelle mondiale ainsi qu’une augmentation de la fréquence des épidémies, et favoriser l’émergence de souches d’entérovirus dans de nouvelles régions géographiques en faisant passer le R0 au-delà du seuil de transmission soutenue de personne à personne. En effet, avec l’émergence récente de la paralysie flasque aiguë due aux entérovirus non poliomyélitiques en Amérique du Nord, y compris au Canada, il est possible que nous assistions actuellement à un phénomène semblable en tout point (Hassel et coll., 2015; Holm-Hansen et coll., 2016; Suresh et coll., 2018). Les vaccins candidats contre l’entérovirus EV-71, un agent important de la MMPB fortement associé aux maladies neuroinvasives, sont en cours de développement; ils ne sont toutefois pas encore disponibles pour utilisation en Amérique du Nord (Zhu et coll., 2014).
Contrairement aux entérovirus, les norovirus et les rotavirus causent la gastroentérite hivernale en Amérique du Nord (Greer et coll., 2009; Patel et coll., 2013). En Espagne, le temps exceptionnellement froid a été associé à un risque accru d’hospitalisations attribuables au rotavirus (Morral-Puigmal et coll., 2018). Bien que le rotavirus soit hautement transmissible de personne à personne, les réservoirs environnementaux (en particulier l’eau) peuvent être importants pour la propagation, et la baisse de la température de l’eau retarde la désintégration du virus dans l’environnement (voir le chapitre 7 : Qualité, quantité et sécurité de l’eau). Des conditions froides et sèches ont également été associées à l’activité des norovirus en Angleterre et au pays de Galles (Lopman et coll., 2009). Par conséquent, le réchauffement climatique au Canada pourrait réduire l’incidence de certains de ces agents pathogènes viraux.
6.3.4
Maladies infectieuses acquises par inhalation à partir de sources environnementales
Partager
Les maladies infectieuses les plus connues chez les humains, qui ont des sources environnementales autres que les humains infectés, les réservoirs d’animaux ou les arthropodes vecteurs, sont la légionellose et la cryptococcose. Ces types d’infections sont causés par des bactéries et des champignons qui persistent dans les sols et l’eau et qui infectent les humains par inhalation après avoir été aérosolisés ou aéroportés dans la poussière. Pour toutes ces infections, bien qu’il y ait eu des études visant à déterminer les effets des conditions météorologiques et climatiques sur leur occurrence, peu d’études ont été menées pour évaluer les impacts potentiels des changements climatiques. Le texte principal fournit des détails sur les agents pathogènes, les maladies qu’ils causent et leur sensibilité aux conditions météorologiques et climatiques, tandis que les effets potentiels des changements climatiques sont décrits dans l’encadré 6.5, l’encadré 6.6, l’encadré 6.7, l’encadré 6.8 et l’encadré 6.9.
6.3.4.1 Légionellose
L’infection par les bactéries Legionella cause la légionellose, qui se présente comme la maladie du légionnaire, la fièvre de Pontiac ou une infection asymptomatique. La maladie du légionnaire se manifeste par une pneumonie (Stout et Yu, 1997), est souvent grave et peut se transformer en insuffisance respiratoire et défaillance de multiples organes avec un taux de mortalité de 11 % à 25 % (Marston et coll., 1994; Stout et Yu, 1997). La fièvre de Pontiac est plus légère et se caractérise par la fièvre, la fatigue, la myalgie et les maux de tête, avec ou sans toux (Tossa et coll., 2006), et les patients se rétablissent après deux à cinq jours sans traitement. Les personnes à risque accru de légionellose comprennent les personnes âgées, les hommes, les fumeurs et celles qui ont des problèmes sous-jacents comme l’immunosuppression, une maladie pulmonaire chronique, le diabète et le cancer (Marston et coll., 1994; Stout et Yu, 1997).
Les bactéries Legionella sont présentes naturellement dans l’eau douce et le sol et se multiplient dans les protozoaires présents dans l’environnement. Elles deviennent un risque pour la santé humaine lorsqu’elles se multiplient dans l’eau entre 25 °C et 42 °C et qu’elles sont aérosolisées et inhalées dans les poumons (Fields et coll., 2002; OMS, 2007). L’infection par Legionella pneumophila (qui cause plus de 70 % des infections humaines) est contractée par inhalation d’aérosols provenant de systèmes d’eau douce dans les maisons ou dans la collectivité (p. ex., tours de refroidissement, spas, pommes de douche, fontaines) (OMS, 2007). La transmission se produit également dans les établissements de soins en raison d’une combinaison de facteurs de risque présents chez les résidents ou les patients et d’une conception et d’un entretien inadéquats de l’infrastructure du bâtiment (Silk et coll., 2013). L’infection de certaines espèces se produit par contact direct ou par aérosolisation de sols ou de compost contaminés (OMS, 2007; Picard-Masson et coll., 2016).
En tant que bactérie présente dans l’environnement, la Legionella est sensible aux facteurs climatiques. La légionellose montre une forte saisonnalité, avec une incidence plus élevée à la fin de l’été et au début de l’automne (Simmering et coll., 2017; Alarcon Falconi et coll., 2018). On observe également des différences régionales, qui peuvent être associées à des facteurs climatiques (Beauté et coll., 2016; Simmering et coll., 2017). L’association avec des facteurs climatiques la plus constante et la plus forte a été signalée entre la légionellose et les précipitations. Plusieurs études menées en Amérique du Nord, en Asie et en Europe ont révélé un accroissement du risque de légionellose sporadique à la suite de précipitations de pluie (Fisman et coll., 2005; Hicks et coll., 2007; Garcia-Vidal et coll., 2013). Le risque est plus grand de deux à onze jours après la pluie (Fisman et coll., 2005; Chen et coll., 2014b; Beauté et coll., 2016;) et suit un rapport de multiplication de 2,5 à 2,6 fois l’incidence pour chaque 5 mm à 1 cm de pluie (Hicks et coll., 2007; (Chen et coll., 2014b) ou de 2,1 à 2,5 fois comparativement aux périodes sans pluie (Fisman et coll., 2005; Beauté et coll., 2016). Cela peut s’expliquer par le fait que les précipitations peuvent augmenter le ruissellement qui entraîne des nutriments et des protozoaires dans les sources d’eau, ce qui peut accroître la réplication des bactéries Legionella en plus d’augmenter la turbidité et la quantité de matière organique et réduire ainsi l’efficacité de la désinfection de l’eau (Fisman et coll., 2005).
Dans les zones tempérées, on a également constaté que les températures plus chaudes augmentaient le risque de légionellose (Fisman et coll., 2005; Hicks et coll., 2007; Conza et coll., 2013; Halsby et coll., 2014; Beauté et coll., 2016; Simmering et coll., 2017). Une hausse de 1 °C a été associée à une augmentation du risque de 2,8 % à 7 % (Fisman et coll., 2005; Hicks et coll., 2007; Conza et coll., 2013), , et le risque survient après un décalage de trois à neuf semaines (Halsby et coll., 2014; Beauté et coll., 2016). Cependant, une étude (Beauté et coll., 2016) a montré que le risque pourrait diminuer à des températures plus élevées (supérieures à 20 °C). Enfin, une humidité relative et une pression de vapeur plus élevées ont été associées au risque de légionellose (Fisman et coll., 2005; Conza et coll., 2013; Gleason et coll., 2016; Simmering et coll., 2017). Une humidité plus élevée a été associée positivement aux cas de légionellose (Gleason et coll., 2016). Simmering et coll. (2017) ont révélé que la température et l’humidité interagissent; le risque le plus élevé se situait entre 16 °C et 27 °C, avec une humidité relative supérieure à 70 %. En résumé, le risque de légionellose semble augmenter après un temps plus chaud et plus humide (Chen et coll., 2014a; Halsby et coll., 2014), car les milieux chauds et humides favorisent la croissance des bactéries Legionella et son aérosolisation.
6.3.4.2 Infection à Cryptococcus gattii
Le Cryptococcus gattii est un champignon présent dans l’environnement qui peut infecter les humains ainsi que de nombreuses espèces animales (Chen et coll., 2014b). Il se trouve dans le sol et les débris d’arbres, mais il peut aussi se trouver dans l’air, l’eau douce et l’eau salée, de même que sur les véhicules et les fomites – des objets ou des matériaux comme des vêtements, des ustensiles et des meubles qui sont susceptibles de propager une infection (Kidd et coll., 2007a). Ses spores présentes au sol ou sur les arbres sont aérosolisées et transportées vers d’autres sites ou inhalées par les humains et les animaux (Kidd et coll., 2007b). Chez les humains, les spores inhalées se retrouvent dans les poumons où elles causent une pneumonie ou des nodules pulmonaires. Elles peuvent être transportées dans la circulation sanguine, en particulier vers le système nerveux central où elles causent une méningite ou des nodules cérébraux (Chen et coll., 2014b) (figure 6.7). Le taux de mortalité est élevé, à savoir 23 % (Phillips et coll., 2015). Les personnes les plus à risque d’infection comprennent les personnes âgées de plus de 50 ans, les fumeurs et les personnes atteintes d’une maladie pulmonaire chronique, de même que celles qui sont immunodéprimées ou qui suivent un traitement immunosuppresseur (MacDougall et coll., 2011).
Cryptococcus gattii vit sous forme de levure dans l’environnement (1), habituellement en association avec certains arbres ou sols autour des arbres. Les humains et les animaux peuvent être infectés par C. gattii après avoir inhalé des cellules de levure ou des spores déshydratées en suspension dans l’air (2), qui traversent les voies respiratoires et pénètrent dans les poumons de l’hôte (3). En raison de leur petite taille, la levure et les spores peuvent se loger profondément dans le tissu pulmonaire. L’environnement à l’intérieur du corps hôte signale à C. gattii de se transformer en sa forme de levure, et les cellules développent d’épaisses capsules en vue de se protéger (4). Les levures se divisent et se multiplient ensuite par bourgeonnement. Après avoir infecté les poumons, les cellules de C. gattii peuvent circuler dans le sang (5) — seules ou dans les cellules macrophages — et infecter d’autres régions du corps, généralement le système nerveux central (6).
Source
CDC des États-Unis, 2020.
Le Cryptococcus gattii est présent dans le monde entier. Il est apparu pour la première fois au Canada en 1999, sur l’île de Vancouver, lorsqu’il a causé une éclosion chez des animaux et des humains en bonne santé (Fyfe et coll., 2008). Depuis, son incidence a augmenté pour atteindre un état stable de 0,2 à 0,5 cas pour 100 000 habitants en Colombie-Britannique (BCCDC, 2019). Depuis 2004, le champignon est présent dans l’environnement et chez les humains et les animaux dans la partie continentale de la Colombie-Britannique et dans les États voisins de Washington et de l’Oregon (MacDougall et coll., 2007). On ne sait pas trop si l’aire de répartition du C. gattii s’est élargie ou s’il était déjà présent dans ces régions (Roe et coll., 2018). Très peu de cas canadiens acquis localement ont été signalés à l’extérieur de la Colombie-Britannique, à l’exception de deux cas chez l’humain au Québec en 2008 et en 2015 (St-Pierre et coll., 2018) et d’un cas dans un chevreuil en Nouvelle-Écosse en 2014 (Overy et coll., 2016).
En tant que champignon présent dans l’environnement, le C. gattii est sensible aux facteurs climatiques. L’humidité, la température, les précipitations, l’évaporation et le rayonnement solaire influent tous sur la présence du C. gattii dans l’environnement (Granados et Castañeda, 2006; Kidd et coll., 2007b). Un modèle de niche écologique utilisé en Colombie-Britannique décrit les zones dont les conditions environnementales favorisent l’établissement du C. gattii comme étant celles de faible altitude, ayant une température quotidienne moyenne supérieure à 0 °C en janvier, et situées dans deux zones climatiques côtières de la Colombie-Britannique où les hivers sont doux et humides et les étés sont chauds et secs (Mak et coll., 2010). Les données d’échantillonnage environnemental en Colombie-Britannique ont révélé que les concentrations du C. gattii dans les échantillons d’air étaient plus élevées durant les mois d’été, lorsque les conditions sont chaudes et sèches et que l’humidité relative est faible (Kidd et coll., 2007b).
La réaction aux facteurs climatiques et à d’autres facteurs environnementaux varie à l’échelle mondiale et peut dépendre en partie de l’endroit où l’on prélève des échantillons du C. gattii (c.-à-d. l’air, le sol ou les arbres), du type de souche et d’autres facteurs. Le vent augmente l’aérosolisation du sol et favorise la propagation (Uejio et coll., 2015). Les chutes de pluie abondantes ou l’humidité peuvent causer le dépôt des spores présentes dans l’air (Kidd et coll., 2007b), bien que, dans la plupart des régions, le C. gattii soit plus susceptible d’être isolé des arbres pendant la saison des pluies, ce qui porte à croire qu’il est plus abondant à ce moment-là (Uejio et coll., 2015). En Colombie-Britannique, la quantité de C. gattii isolé à partir des arbres et sa concentration dans le sol sont plus faibles lorsque les températures sont plus élevées, mais dans d’autres parties du monde, la fréquence d’isolement était la plus forte à des températures très élevées (Uejio et coll., 2015). Les spores de C. gattii peuvent résister à un fort rayonnement solaire, de sorte qu’il est peu probable que les conditions estivales avec des températures élevées et un taux d’isolement élevé empêchent le C. gattii de survivre (Rosas et Casadevall, 2006).
6.3.4.3 Blastomycose
Le Blastomyces dermatitidis est un champignon dimorphique présent dans le sol humide et la végétation en décomposition. La transmission se produit de l’environnement aux humains et aux autres animaux par aérosolisation et inhalation (Castillo et coll., 2016). Les spores inhalées dans les poumons causent une blastomycose pulmonaire caractérisée par la toux, la fièvre et l’essoufflement (McBride et coll., 2017). La levure peut être transportée par le sang vers d’autres organes et l’infection cutanée peut se produire par inoculation directe (Castillo et coll., 2016). Le taux de mortalité est élevé (entre 5 % et 20 %), probablement en raison d’un diagnostic tardif, en particulier dans les régions moins endémiques (Crampton et coll., 2002; Dalcin et Ahmed, 2015). L’infection est traitée avec des antifongiques (Castillo et coll., 2016). La période d’incubation est habituellement d’un à trois mois, et les cas surviennent principalement chez les personnes immunodéprimées, les fumeurs et les personnes atteintes de maladies sous-jacentes qui présentent un risque plus élevé d’infection et de mortalité (Crampton et coll., 2002; Dalcin et Ahmed, 2015; Castillo et coll., 2016; McBride et coll., 2017). Les personnes sont plus à risque d’être exposées pendant les activités de plein air comme la chasse et le camping (Crampton et coll., 2002). La plupart des cas sont sporadiques, mais les activités qui perturbent le sol (p. ex., construction, déforestation, activités de plein air) et l’exposition aux berges de rivière ont été associées à des éclosions de sources ponctuelles (Baumgardner et Burdick, 1991; Proctor et coll., 2002; Azar et coll., 2015).
Le Blastomyces dermatitidis est endémique dans certaines régions du Canada, dont le sud de la Saskatchewan, le Manitoba, l’Ontario et le sud du Québec (Crampton et coll., 2002; Litvinov et coll., 2013; Lohrenz et coll., 2018). La blastomycose n’est à déclaration obligatoire qu’en Ontario et au Manitoba et peut être sous-estimée dans d’autres régions du pays. Il est hyperendémique dans le nord-ouest de l’Ontario, caractérisé par des taux élevés et persistants de maladie. La région de Kenora a les taux les plus élevés déclarés dans le monde, avec 35 hospitalisations par 100 000 habitants par année dans le nord-ouest de l’Ontario (Litvinjenko et Lunny, 2017; Brown et coll., 2018). Cette région endémique est contiguë aux régions endémiques des États américains qui bordent les rivières Mississippi et Ohio, les Grands Lacs et la voie maritime du Saint-Laurent (McBride et coll., 2017).
Le Blastomyces est un mycète présent dans l’environnement; sa croissance et sa dispersion dépendent donc des conditions climatiques. Les facteurs écologiques particuliers sont mal compris en partie parce que l’organisme a rarement été récupéré de l’environnement, mais le sol humide près des cours d’eau et des lacs semble être la niche privilégiée des mycètes de l’espèce Blastomyces (Castillo et coll., 2016). La perturbation du sol facilite l’aérosolisation (Baumgardner, 1997). On suppose que la proximité des cours d’eau et la sécheresse suivie d’une augmentation des précipitations peuvent contribuer à l’exposition des spores et faciliter leur dispersion (McDonough et coll., 1976; McTaggart et coll., 2016). Bien que le pic d’apparition de l’infection pulmonaire se situe à l’automne et à l’hiver, en raison de l’exposition pendant le temps plus chaud (Morris et coll., 2006; Light et coll., 2008; Dalcin et Ahmed, 2015; Brown et coll., 2018), peu d’études ont évalué expressément le rôle des facteurs climatiques, et des résultats variables ont été rapportés. Baumgardner et coll. (2011) ont noté un lien entre les précipitations totales, la température (températures moyennes basses pendant la saison de l’infection, mais températures maximales élevées la saison précédente) et les cas chez les canidés. Seitz et coll. (2015) ont relevé un risque plus élevé d’hospitalisations pour blastomycose lorsque la température maximale est plus basse. Proctor et coll. (2002) ont enquêté sur une éclosion qui a suivi une période de sécheresse prolongée. De même, Pfister et coll. (2011) ont déterminé qu’une sécheresse modérée à grave, suivie de précipitations supérieures à la normale et d’un réchauffement des déchets de jardin ont été associés à une éclosion au Wisconsin. Le vent peut être un facteur, car certaines éclosions ont été associées à la poussière d’excavation (Baumgardner et coll., 2011) ou à la perturbation des déchets de jardin en décomposition (Pfister et coll., 2011). Un modèle de niche écologique du Wisconsin a révélé que l’indice de végétation estivale et la proximité des voies navigables étaient les meilleurs facteurs prédictifs de l’emplacement des cas de blastomycose humaine et animale, et que les facteurs bioclimatiques ne jouaient pas un rôle important (Reed et coll., 2008).
6.3.4.4 Histoplasmose
L’histoplasmose est causée par le champignon dimorphique Histoplasma capsulatum. Les spores prolifèrent dans un sol humide riche en guano de chauves-souris ou d’oiseaux le long des cours d’eau et dans les grottes. La perturbation du sol attribuable aux travaux d’excavation et de construction entraîne la production d’aérosols et un risque d’inhalation de spores. L’infection se développe de 7 à 14 jours après l’exposition. Elle est souvent asymptomatique ou cause des maladies spontanément résolutives avec fièvre, maux de tête, faiblesse, douleurs thoraciques et toux. Elle peut engendrer une infection extrapulmonaire et aussi causer une infection pulmonaire plus grave chez les personnes atteintes de maladies pulmonaires sous-jacentes ou immunodéprimées (Kauffman, 2007). L’Histoplasma capsulatum est endémique le long des bassins hydrographiques du Saint-Laurent et des Grands Lacs au Québec et en Ontario, dans les vallées de l’Ohio et du Mississippi aux États-Unis, et le long des voies navigables en Asie et en Amérique du Sud. Au Canada, c’est au Québec que l’on note le taux le plus élevé de maladies diagnostiquées. Dans une étude, on a recensé 58 cas au Canada sur une période de trois ans (de 1992 à 1994), 72 % ayant été signalés au Québec (Nicolle et coll., 1998). Des cas en Alberta ont été recensés pour la première fois en 2003, et depuis, on en signale quelques-uns chaque année (Anderson et coll., 2006; Alberta Health, 2018). Le champignon est sensible au climat, préférant des températures modérées (de 18 °C à 28 °C), une humidité constante et une faible lumière (Teixeira, 2016).
6.3.4.5 Coccidioïdomycose
La coccidioïdomycose est causée par les mycètes de l’espèce Coccidioides que l’on trouve dans le sol aride, y compris le C. immitis en Californie et le C. posadasii ailleurs (Maves et Crum-Cianflone, 2012). Les spores sont libérées dans l’air lorsque le sol est perturbé et peuvent être inhalées par les humains, causant la coccidioïdomycose, aussi connue sous le nom de fièvre de la vallée. La maladie a une période d’incubation d’une à trois semaines, et de nombreuses personnes demeurent asymptomatiques ou ne présentent que de légers symptômes. Environ 40 % développent une infection pulmonaire accompagnée de toux, de douleurs thoraciques, de dyspnée et de fièvre. Cette situation est habituellement spontanément résolutive et la personne guérit en quelques semaines. L’infection disséminée se produit chez 5 % des personnes et entraîne une maladie pulmonaire diffuse et chronique, une méningite ou une infection dans d’autres organes (Maves et Crum-Cianflone, 2012). Cette forme grave nécessite un traitement antifongique prolongé. Les femmes enceintes et les personnes dont l’immunité cellulaire est affaiblie, comme en raison d’une séropositivité au VIH ou d’une transplantation d’organes, sont plus à risque de développer une maladie grave. La réactivation de la maladie latente est également possible (Maves et Crum-Cianflone, 2012). L’aire de répartition géographique du mycète comprend des régions de l’Amérique du Sud et de l’Amérique centrale, le Mexique et le sud-ouest des États-Unis (Maves et Crum-Cianflone, 2012). Certaines régions de la Californie et de l’Arizona affichent les taux les plus élevés au monde. La perturbation des sols, y compris la construction, les fouilles archéologiques, les tremblements de terre et les tempêtes de vent, dans les zones désertiques endémiques sont la principale source d’exposition (Maves et Crum-Cianflone, 2012).
Le Coccidioides spp. est affecté par des facteurs climatiques (Nguyen et coll., 2013); il a besoin d’humidité pour proliférer dans le sol, puis d’une période sèche pour produire des spores pouvant passer à l’état d’aérosol (l’hypothèse « croissance et dispersion ») (Comrie et Glueck, 2007). Le pic saisonnier des cas chez l’humain aux États-Unis se produit principalement à l’automne, après une période de mois pluvieux suivie de mois chauds et secs. On observe également une tendance cyclique avec des pics d’incidence environ tous les deux à cinq ans qui seraient associés à des sécheresses suivies de temps pluvieux (Park et coll., 2005; Tamerius et Comrie, 2011; Gorris et coll., 2018 ). Les températures plus chaudes sont associées à une incidence plus élevée chez les humains (Gorris et coll., 2018), et un accroissement de l’incidence est observé à la suite de grandes tempêtes de vent (Tong et coll., 2017). Cependant, l’incidence est faible dans les régions où les précipitations annuelles moyennes sont élevées ou où le sol est très humide (comme les régions côtières) (Gorris et coll., 2018).
6.4
Adaptation en vue de réduire les risques pour la santé
Partager
Le Canada dispose déjà d’un solide système de santé publique, qui est relié aux organisations internationales de santé publique. Ensemble, ces systèmes contribuent à la préparation aux maladies infectieuses, à la surveillance, au suivi et à l’intervention en cas d’éclosion. Comme les risques de maladies infectieuses continuent de croître dans un climat en évolution, ces activités seront de plus en plus importantes pour le succès de l’adaptation. La préparation en amont et l’intervention en cas d’éclosion sont deux volets des activités de santé publique dans le contexte des maladies infectieuses émergentes et réémergentes. Ces volets comportent trois activités de santé publique principales, soit : l’évaluation des risques en vue de cerner les risques actuels et futurs; la surveillance des risques connus ou des éventuels risques émergents cernés dans les évaluations des risques; et les interventions visant à prévenir et à contrôler les maladies infectieuses. Ces trois activités sont examinées individuellement, mais en pratique, elles devraient être étroitement liées; les évaluations des risques guident la mise en œuvre rationnelle des activités de surveillance qui, à leur tour, déclenchent des interventions pour protéger le public lorsqu’un aléa est détecté (figure 6.8). Les mesures visant à protéger la santé doivent être robustes et coordonnées pour contrer les risques accrus de maladies infectieuses imputables aux changements climatiques. De plus, étant donné que bon nombre des maladies infectieuses émergentes ou réémergentes impliquent des zoonoses transmises directement ou par vecteur, l’approche « Un monde, une santé » (aussi connu sous le nom d’Une seule santé), qui tient compte des interactions entre les facteurs humains, animaux et environnementaux dans la transmission, l’émergence et la réémergence des maladies, est nécessaire à l’élaboration d’interventions efficaces (Ogden et coll., 2017; Ogden et coll., 2019).
En général, le Canada et d’autres pays à revenu élevé pourraient être considérés comme bien placés pour réagir aux maladies infectieuses émergentes et réémergentes grâce à de solides institutions internationales de surveillance et d’intervention, telles que : le Règlement sanitaire international (RSI) de l’OMS (OMS, 2019c); le Réseau mondial d’alerte et d’action en cas d’épidémie (OMS, 2019d); et le système de soins de santé et les institutions de santé publique au Canada. Le système de santé publique canadien s’articule autour du partenariat fédéral-provincial-territorial qu’est le Réseau pancanadien de santé publique (Réseau pancanadien de santé publique, 2016), qui est coordonné par l’ASPC du gouvernement fédéral. Ce système est appuyé par des établissements d’enseignement et des entreprises de grande qualité au Canada qui peuvent faciliter l’innovation et les réponses aux menaces des maladies infectieuses émergentes. Le Canada est un chef de file pour ce qui est de l’approche « Un monde, une santé » (ou « Une seule santé ») dans les établissements fédéraux, provinciaux et universitaires, et cette approche est mise en œuvre de façon systématique pour composer avec ces menaces (gouvernement du Canada, 2013; INSPQ, 2018a; CPHAZ, 2019; GREZOSP, 2019; Ogden et coll., 2019). En dépit de cette capacité, il y a beaucoup d’inconnues à l’horizon et, comme il a été constaté avec la COVID-19, même les systèmes de santé publique les mieux préparés peuvent être se retrouver dans une position très précaire en raison des maladies infectieuses émergentes.
Deux volets des mesures de santé publique visant à gérer les maladies infectieuses émergentes et réémergentes.
Figure 6.8
Deux volets des mesures de santé publique visant à gérer les maladies infectieuses émergentes et réémergentes. Les boîtes bleues indiquent une intervention en cas d’éclosion et les boîtes oranges représentent l’état de préparation en amont. Des exemples d’activités qui peuvent être des composantes de la gestion de l’éclosion sont présentés dans les cases de couleur vert. Les lignes pointillées indiquent comment l’intervention en cas d’éclosion peut être déclenchée par la détection précoce du risque de maladie au moyen d’une approche de préparation en amont.
Depuis la dernière évaluation nationale des changements climatiques et de la santé au Canada (Berry et coll., 2014a), l’ASPC a amélioré la préparation en amont pour les risques émergents et réémergents de maladies à transmission vectorielle grâce à un programme intégré d’adaptation qui comprend les éléments suivants (figure 6.8) :
le classement par priorité des risques de maladie devant faire l’objet d’études, maintenant officialisé dans un processus d’analyse décisionnelle multicritères (ADMC) (Ng et coll., 2019);
des études de modélisation visant à définir les liens entre les risques liés au climat et aux maladies et à prévoir où et quand les changements climatiques pourraient conduire à l’émergence de maladies (Ng et coll., 2019);
la mise en œuvre de programmes de surveillance pour valider les modèles et suivre les maladies à transmission vectorielle émergentes (Drebot, 2015; Ogden et coll., 2014c);
la synthèse des connaissances (Hierlihy et coll., 2019), des outils de prévision des maladies (Ripoche et coll., 2019) et de communication des risques (figure 6.9) pour appuyer les efforts déployés à l’échelle locale en vue de l’adaptation aux risques croissants des maladies à transmission vectorielle.
6.4.1
Évaluation des risques en vue de cerner les risques actuels et futurs
Partager
6.4.1.1 Évaluations rapides et réactives du risque
Les évaluations rapides et réactives du risque sont des évaluations qualitatives du risque réalisées en réponse à un événement immédiat d’émergence d’une maladie au Canada, ou à un risque perçu, comme un événement ou une menace de maladie internationale, qui nécessite une prise de décision rapide. Elles sont élaborées à l’aide de la littérature scientifique et des connaissances sur une maladie au moment où la maladie émerge ou réapparaît; elles peuvent être mises à jour à mesure que l’on en sait davantage au sujet de l’événement d’émergence de la maladie. Elles précèdent généralement la mise en œuvre des activités de surveillance et d’intervention et visent à les orienter. Les exemples comprennent celles menées en réponse à la menace du virus Zika au Canada aux échelles nationale (gouvernement du Canada, 2016) et provinciale (INSPQ, 2016), de même que celles qui ont été élaborées par le Centre européen de prévention et de contrôle des maladies (ECDC) en réponse à un certain nombre d’événements d’émergence de maladies (ECDC, 2019). L’Organisation mondiale de la Santé a élaboré un protocole pour la préparation liée à ces évaluations (OMS, 2019e) au Canada. Il est essentiel que les responsables canadiens de la santé publique puissent effectuer des évaluations rapides et réactives des risques pour pouvoir intervenir en cas d’éclosion, y compris celles causées par les changements climatiques.
6.4.1.2 Prévision des populations à risque futures et élaboration d’alerte précoce au moyen d’évaluations quantitatives des risques fondées sur des modèles
Les évaluations quantitatives des risques visent à prédire où et quand les maladies infectieuses peuvent émerger ou réapparaître avec une précision utile aux responsables de la santé publique afin qu’ils puissent adopter les politiques et programmes nécessaires pour se préparer et intervenir face à l’émergence de maladies. Les évaluations des risques peuvent utiliser des modèles mathématiques qui recréent les composantes essentielles des cycles de vie des vecteurs ou des cycles de transmission des maladies, s’il y a suffisamment de données quantitatives pour calibrer ces types de modèles. Sinon, des modèles de niche statistique ou écologique peuvent être utilisés, à condition qu’il y ait des données de surveillance comprenant à tout le moins des renseignements sur la présence, mais préférablement des données sur la présence, l’absence et l’abondance pour les calibrer. Dans un cas comme dans l’autre, l’objectif des modèles est d’évaluer où et quand le risque de maladies infectieuses sensibles au climat peut changer (c.-à-d. qu’ils fournissent une « alerte précoce » des risques futurs de maladie). Les modèles prédictifs sont utilisés à trois fins principales (Ogden et Gachon, 2019) :
la prévision à court terme des risques de maladie en fonction des conditions météorologiques;
les prévisions à long terme ou saisonnières fondées sur le climat;
les projections des effets à long terme des changements climatiques sur les risques de maladie.
Les modèles sont élaborés en établissant d’abord les associations clés entre la présence de vecteurs ou d’agents pathogènes et les conditions météorologiques (pour les prévisions à court terme) ou les variables climatiques, tout en tenant compte d’importants déterminants de leur présence non liés au climat qui peuvent comprendre d’autres changements environnementaux tels que le changement dans l’aménagement du territoire (Patz et coll., 2003). La validation des associations déduites par rapport aux données réelles est importante pour donner confiance dans l’utilisation des produits du modèle. Une fois que les associations entre le climat et la présence d’agents pathogènes/vecteurs ont été établies et quantifiées, il est possible d’évaluer les impacts des changements climatiques anticipés, obtenus à partir de modèles climatiques régionaux et mondiaux, sur la répartition d’agents pathogènes/vecteurs et les risques connexes.
Les prévisions fournissent une alerte précoce des éclosions de maladies causées par les conditions météorologiques. Cela peut comprendre des prévisions à court terme s’échelonnant sur quelques jours ou semaines et des prévisions à long terme s’échelonnant sur plusieurs mois qui peuvent être utilisées pour mettre en œuvre des activités de prévention et d’intervention (Morin et coll., 2018). Les évaluations fondées sur des modèles des effets des changements climatiques fournissent des évaluations de l’endroit et du moment où les événements d’émergence et de réémergence de maladies pourraient se produire dans les décennies à venir, et sont utilisées dans un certain nombre de contextes (Ogden et coll., 2014c), dont :
la prise de décisions stratégiques nationales de haut niveau sur la nécessité pour la santé publique de se préparer à de nouveaux risques (Centre des maladies infectieuses d’origine alimentaire, environnementale et zoonotique, 2017);
les évaluations provinciales, territoriales et municipales de la vulnérabilité aux impacts des changements climatiques sur la santé et des mesures d’adaptation nécessaires (Berry et coll., 2014b);
la conception et la mise en œuvre de la surveillance (Ogden et coll., 2014).
Comme nous l’avons vu plus haut dans le présent chapitre, un certain nombre d’évaluations des risques fondées sur des modèles ont été réalisées pour le Canada, y compris pour la maladie de Lyme (Ogden et coll., 2006; Ogden et coll., 2014)), la fièvre récurrente (Sage et coll., 2017), les tiques vectrices de Dermacentor variabilis (Minigan et coll., 2018), des maladies exotiques transmises par les moustiques et leurs vecteurs (Ogden et coll., 2014a; Ng et coll., 2017), et le virus du Nil occidental (Hongoh et coll., 2012; Chen et coll., 2013).
6.4.2
Établissement des priorités
Partager
Depuis que Charron et coll. (2008) et l’OMS (OMS, 2019g) ont déterminé qu’il s’agissait d’un besoin méthodologique important, un certain nombre de méthodes d’analyse décisionnelle multicritères (ADMC) ont été explorées afin d’établir l’ordre de priorité des maladies à étudier en fonction de leur importance sur le plan de la santé publique, y compris leur risque d’émergence ou de réémergence en raison des changements climatiques (Ng et Sargeant, 2012; Cox et coll., 2013). Ces types de méthodes ont été adoptés et parfois adaptés pour l’établissement des priorités en matière de maladies aux échelons fédéral et provincial (INSPQ, 2018b), mais pas encore par toutes les administrations canadiennes.
6.4.3
Surveillance des risques connus ou des risques futurs possibles
Partager
Deux types de surveillance sont utilisés pour déceler les maladies infectieuses émergentes afin d’éclairer la gestion des éclosions. Le premier est la vigilance à l’égard des événements d’émergence de maladies à l’échelle internationale, et le deuxième, la surveillance propre à une maladie qui est mise en œuvre une fois qu’une maladie est reconnue comme présentant un risque important pour la santé publique de la population canadienne. L’impact possible des changements climatiques sur l’augmentation des événements d’émergence et de réémergence des maladies infectieuses à l’échelle internationale, et sur la dispersion croissante des agents pathogènes par les voyages, le commerce et la migration, accroît la nécessité d’une vigilance efficace à l’égard des maladies infectieuses émergentes dans d’autres pays. Les efforts internationaux qui soutiennent ce type de surveillance comprennent le RSI, aux termes duquel tous les États membres de l’OMS sont tenus de surveiller les événements d’émergence de maladies et de signaler tout cas repéré. En outre, dans le cadre de l’initiative mondiale « Un monde, une santé » (ou « Une seule santé »), l’Organisation des Nations Unies pour l’alimentation et l’agriculture, l’Organisation mondiale de la santé animale et l’OMS collaborent au Système mondial d’alerte précoce, qui est un système d’alerte pour les menaces de maladies émergentes à l’interface homme-animal-écosystèmes. Les programmes internationaux de surveillance « passive » comprennent le réseau de surveillance GeoSentinel, un réseau international de cliniques médicales qui participent sur une base volontaire, et des systèmes de surveillance qui examinent le contenu Web accessible au public pour détecter les signaux de menaces émergentes potentielles. Il s’agit notamment du Program for Monitoring Emerging Diseases (ProMED), de HealthMap et de Medisys, de même que du Réseau mondial d’information sur la santé publique (RMISP) mis sur pied par l’ASPC (Ogden et coll., 2017).
Au Canada, un certain nombre de méthodes sont disponibles et sont utilisées pour assurer la surveillance des maladies infectieuses endémiques et émergentes, afin de suivre les tendances au chapitre de leur incidence, de déterminer leur répartition spatiotemporelle et la propagation des cas (y compris les grappes) et de définir les facteurs de risque au sein de la population touchée. La méthode normalisée pour obtenir des données sur les cas de maladies humaines à l’échelle nationale est le Système canadien de surveillance des maladies à déclaration obligatoire (SCSMDO) coordonné par l’ASPC (gouvernement du Canada, 2019e). Certaines maladies qui ne sont pas à déclaration obligatoire à l’échelle nationale peuvent l’être aux échelons provincial ou territorial. Ces maladies à déclaration obligatoire sont repérées à l’aide de critères de définition de cas cliniques et de laboratoire, notamment l’émergence ou la réémergence (à l’échelon national ou international). Parmi les autres critères pour l’ajout des maladies à la liste, mentionnons les maladies sous surveillance internationale, l’incidence au Canada, la gravité, la transmissibilité, la possibilité d’éclosions, le fardeau socioéconomique, la capacité de prévention, la perception du risque et la nécessité d’intervention de la santé publique (Agence de la santé publique du Canada, 2009).
D’autres méthodes de collecte d’information sur les cas possibles de maladies humaines pendant les événements d’émergence comprennent la surveillance syndromique, par exemple, l’utilisation de données en temps réel des pharmacies sur les ventes de remèdes contre la grippe pour suivre l’évolution de la pandémie de grippe H1N1 au Canada en 2009 (Muchaal et coll., 2015). Une autre méthode est la surveillance passive en laboratoire. En réponse à l’accroissement possible des risques posés par les arbovirus au Canada, le Laboratoire national de microbiologie de l’ASPC a mis en place une surveillance des cas humains d’infection par un arbovirus autre que le VNO en procédant régulièrement au dépistage d’un panel d’arbovirus endémiques et exotiques dans les échantillons de sérum prélevés aux fins de dépistage du VNO qui se sont avérés négatifs pour celui-ci (Drebot, 2015). Le Réseau pancanadien de santé publique a la capacité d’instaurer une surveillance nationale des cas humains pendant des périodes limitées lors d’événements d’émergence de maladies. Ce fut le cas récemment en réponse à la pandémie de COVID-19 et à l’éclosion du virus Zika en Amérique centrale et en Amérique du Sud en 2015-2016, qui a infecté plus de 500 voyageurs canadiens (gouvernement du Canada, 2019f).
Beaucoup d’efforts ont été déployés au Canada pour utiliser la surveillance sentinelle des animaux et des vecteurs afin de détecter les risques liés aux zoonoses à transmission vectorielle émergentes, en particulier le VNO, la maladie de Lyme et d’autres agents pathogènes transmis par l’Ixodes spp. comme méthode d’alerte précoce de la propagation des maladies émergentes et, dans le cas du VNO, des éclosions imminentes (Thomas-Bachli et coll., 2015). Quant aux maladies à transmission vectorielle, la surveillance des vecteurs comprend une surveillance active pour capturer les moustiques vecteurs, qui sont ensuite analysés pour savoir s’ils sont porteurs d’agents pathogènes à transmission par les moustiques, en particulier le VNO. Ce type de surveillance est réalisé couramment dans de nombreuses administrations au Canada (gouvernement du Canada, 2019a). Elle comprend également la collecte de tiques par échantillonnage par filet traînant, la capture d’hôtes de tiques et les analyses de tiques et de leurs hôtes pour détecter la présence d’agents pathogènes transmis par les tiques (Ogden et coll., 2014). La surveillance passive des tiques, qui englobe la présentation de tiques par les patients de cliniques médicales et vétérinaires, est une activité de surveillance clé au Canada, fournissant un ensemble de données particulièrement important qui vient étayer l’attribution de l’expansion de l’aire de répartition de I. scapularis au Canada aux impacts des changements climatiques (Leighton et coll., 2012).
Tant pour les moustiques que les tiques, la surveillance des vecteurs permet de capturer de multiples espèces de vecteurs et d’agents pathogènes. La surveillance des moustiques pour détecter le VNO a permis d’identifier des incursions d’Aedes spp. exotiques dans le sud de l’Ontario, qui sont les vecteurs du virus de la dengue et du virus Zika (unité sanitaire du comté de Windsor-Essex, 2019a, unité sanitaire du comté de Windsor-Essex, 2019b). La surveillance des tiques est principalement assurée par deux méthodes : la surveillance passive, qui consiste à analyser les tiques que les membres du grand public ont retirées sur eux-mêmes ou sur leurs animaux de compagnie et ont soumises par l’intermédiaire des cabinets vétérinaires et médicaux participants; et la surveillance active, qui consiste à recueillir des tiques à la recherche d’un hôte en traînant une flanelle sur le sol (échantillonnage par filet traînant) ou en examinant les rongeurs sauvages capturés (Ogden et coll., 2014c). Ces formes de surveillance des tiques ont été conçues à l’origine pour cerner le risque de la maladie de Lyme, mais elles sont capables de détecter le risque d’autres espèces de vecteurs de maladie (Gabriele-Rivet et coll., 2015; Nelder et coll., 2019). Les tiques recueillies dans le cadre des activités de surveillance font l’objet d’un dépistage de routine ciblant un large éventail d’agents pathogènes, en plus de la bactérie causant la maladie de Lyme (Dibernardo et coll., 2014). Au Canada, l’espèce à laquelle appartiennent la plupart des tiques recueillies au moyen de la surveillance est actuellement déterminée dans des laboratoires provinciaux, puis la tique est analysée pour détecter la présence d’agents pathogènes au Laboratoire national de microbiologie. Toutefois, cette méthode ne sera pas viable une fois que les tiques seront plus répandues, compte tenu de la forte augmentation prévue et observée (Gasmi et coll., 2018) du nombre de présentations de tiques au Canada. Il faut adopter des systèmes de surveillance nouveaux et nécessitant moins de ressources pour protéger la population canadienne contre l’accroissement des risques liés à ces maladies.
Un projet de science citoyenne appelé eTick a été lancé en 2014. Il s’agit d’une plateforme Web qui permet aux membres du grand public de soumettre des images de tiques aux fins d’identification de l’espèce par un entomologiste (eTick, 2019). Ce projet élargit considérablement la couverture géographique potentielle de la surveillance des tiques, en plus de donner l’occasion de fournir rapidement des renseignements sur la santé publique directement à la population exposée aux tiques. Il permet d’assurer une surveillance en temps réel de diverses espèces de tiques et de cartographier les lieux où elles sont observées, y compris les changements de répartition. Le programme est actuellement offert au Québec, en Ontario et au Nouveau-Brunswick. Au Manitoba, un système d’identification par image appelé Tick Checker, semblable à eTick, a été mis en œuvre en 2017 (gouvernement du Manitoba, 2019a). Cet effort s’inscrit dans le cadre du programme de surveillance passive des tiques à pattes noires du Manitoba, qui vise à cerner les endroits où de nouvelles populations de tiques et le risque de maladie de Lyme pourraient émerger, avant que ce ne soit confirmé par la surveillance active des tiques (gouvernement du Manitoba, 2018).
Les organismes de santé du bétail, comme l’Agence canadienne d’inspection des aliments, assurent le suivi de certaines zoonoses chez les animaux sentinelles lorsqu’elles relèvent de leur mandat, par exemple, le VEEE chez les chevaux (gouvernement du Canada, 2018c). Les organismes de santé de la faune, comme le Réseau canadien de la santé de la faune, aident à suivre certaines zoonoses d’origine faunique chez les animaux sentinelles (RCSF, 2019).
6.4.4
Interventions visant à prévenir et à contrôler l’incidence des maladies
Partager
Les interventions internationales en cas d’éclosion de maladies infectieuses émergentes sont mandatées en application du RSI par l’intermédiaire du Réseau mondial d’alerte et d’action en cas d’épidémie (GOARN) [OMS, 2019d]. Ce réseau réunit des ressources techniques et opérationnelles d’institutions pertinentes dans les États membres de l’OMS, y compris des initiatives de surveillance, des réseaux de laboratoires, des organisations des Nations Unies (p. ex., UNICEF, HCNUR), la Croix-Rouge et des organisations humanitaires internationales non gouvernementales telles que Médecins sans frontières. Le réseau a la capacité de réunir et de déployer des équipes techniques dans les pays touchés par les éclosions afin d’aider à la coordination, à la gestion des maladies cliniques, à l’analyse épidémiologique ainsi qu’à la logistique et aux communications nécessaires pour lutter contre les éclosions. L’Agence de la santé publique du Canada dirige la vaste participation du Canada au GOARN. Elle a été créée en 2004, en partie pour assurer et améliorer la coordination nationale des interventions en cas de maladies infectieuses émergentes par l’intermédiaire du Réseau pancanadien de santé publique, et pour participer au RSI ainsi qu’au GOARN.
Le système de soins de santé et sa capacité à relever les cas de maladies humaines et de les traiter sont des moyens clés de réduire les risques pour la population canadienne en cas d’éclosion. Le diagnostic et le traitement des cas réduisent la transmission et contribuent à lutter contre les éclosions. Cependant, cela requiert de la vigilance de la part des travailleurs de la santé de première ligne; pendant certaines éclosions, les établissements de santé peuvent devenir un point de convergence pour la transmission, comme cela s’est produit pendant l’éclosion du SRAS au Canada (Varia et coll., 2003). L’accroissement de la résistance aux antimicrobiens et aux antiviraux remet en question la capacité des systèmes de soins de santé à traiter les maladies émergentes (OMS, 2019f).
La capacité technologique du Canada à prévenir et à contrôler les maladies infectieuses est bien avancée. Le Réseau fédéral-provincial-territorial des laboratoires de santé publique du Canada, appuyé par le Laboratoire national de microbiologie de l’ASPC, qui est l’une des sept installations de confinement de niveau 4 en Amérique du Nord, a la capacité de développer, d’améliorer et de mettre en œuvre des tests diagnostiques pour les maladies infectieuses émergentes telles que le virus Zika (Safronetz et coll., 2017). Le réseau mondial d’organisations de santé publique et d’entreprises pharmaceutiques est capable de mettre au point des vaccins en réponse aux éclosions (Carlsen et Glenton, 2016; Henao-Restrepo et coll., 2017). L’OMS a dressé une liste d’agents pathogènes prioritaires pour lesquels des vaccins sont nécessaires de toute urgence; plusieurs de ces maladies sont des zoonoses qui peuvent se propager de personne à personne, et le risque d’émergence peut être accru par les changements climatiques. Toutefois, le temps qu’il faut pour mettre au point des vaccins et pour en évaluer l’innocuité et l’efficacité afin d’assurer leur acceptabilité par le public (Carlsen et Glenton, 2016) souligne la nécessité de s’y prendre à l’avance plutôt que de simplement compter sur la mise au point de vaccins pour gérer les éclosions. Bien qu’il y ait une grande capacité de découverte de vaccins au Canada (Plummer et Jones, 2017), la pandémie de COVID-19 a mis en relief le manque de capacité de fabrication de vaccins au Canada.
D’autres adaptations technologiques sont importantes pour le contrôle actuel et futur des maladies à transmission vectorielle au Canada. Pour les maladies transmises par les moustiques comme le VNO, on a principalement recours aux méthodes de lutte contre les moustiques, qui sont efficaces. Elles consistent notamment à tuer les larves dans leur habitat aquatique (larvicide) à l’aide d’agents de lutte biologique non chimiques comme le Bacillus thuringiensis israelensis et à tuer les moustiques adultes (adulticide) au moyen de pulvérisations chimiques comme la deltaméthrine (Reisen et Brault, 2007). Dans le cas des maladies transmises par les tiques comme la maladie de Lyme, les méthodes de lutte contre les vecteurs, comme la pulvérisation d’acaricides à l’échelle de la région, ne sont pas homologuées et ne sont donc pas disponibles, ou en sont à un stade expérimental de développement au Canada (Ogden et coll., 2015). Les mesures de protection personnelle demeurent les mesures les plus efficaces pour lutter contre les maladies à transmission vectorielle, notamment les insectifuges, qui préviennent les morsures de tiques, les vêtements traités à l’insecticide (perméthrine) et l’élimination rapide des tiques qui se nourrissent sur le corps pour prévenir la transmission du B. burgdorferi (Ogden et coll., 2015). Pour que ces mesures soient efficaces, le public doit comprendre les risques, les percevoir comme méritant qu’on y donne suite, connaître les mesures de protection et être prêt à les prendre. Pour ce faire, il faut des communications efficaces de la part des organismes de santé publique à tous les niveaux, ce qui est de plus en plus difficile parce que la désinformation, surtout sur Internet, est un problème croissant (Greenberg et coll., 2017). En plus de communiquer de l’information sur la protection individuelle contre les maladies au Canada, il sera de plus en plus important de fournir des renseignements aux voyageurs. Cela aidera à prévenir l’introduction des infections à leur retour au Canada, en particulier des maladies exotiques à transmission vectorielle, qui pourraient être de plus en plus susceptibles d’être de transmission autochtone dans notre pays en raison des changements climatiques (Berrang-Ford et coll., 2009).
6.4.5
Défis en matière d’adaptation pour les collectivités et les membres de la société exposés à un risque accru
Partager
La vulnérabilité aux risques de maladies infectieuses liés aux changements climatiques est complexe et déterminée par un certain nombre de facteurs qui interagissent pour modifier l’exposition, la sensibilité et la capacité d’adaptation d’une personne. Bien que tous les Canadiens et les Canadiennes soient vulnérables aux changements climatiques, les impacts et les risques ne sont pas les mêmes pour tous, et certaines personnes et collectivités seront touchées de façon disproportionnée (voir le chapitre 9 : Changements climatiques et équité en santé). Les iniquités existantes en matière de santé et les variations de l’état des déterminants de la santé, combinées à l’exposition (p. ex., emplacement géographique) et aux facteurs génétiques et biologiques, peuvent accroître ou réduire la vulnérabilité aux changements climatiques. Il existe des lacunes au chapitre des connaissances concernant la combinaison de variables qui accroissent la vulnérabilité aux risques de maladies infectieuses liés aux changements climatiques. Cependant, on s’attend à ce que les personnes qui présentent des déterminants d’un mauvais état de santé (p. ex., faible revenu, logement non conforme aux normes, insécurité alimentaire) soient plus susceptibles de ressentir des impacts disproportionnés et d’avoir une capacité limitée à prendre des mesures de protection. Par exemple, en général, un mauvais état de santé est associé à un statut socioéconomique inférieur qui, à son tour, est associé à des taux plus élevés de comportements et d’habitudes alimentaires malsains (Hajizadeh et coll., 2016) engendrant des taux plus élevés de maladies chroniques (Roberts et coll., 2015) et pouvant accroître les risques sanitaires associés aux impacts des changements climatiques sur les maladies infectieuses.
Au Canada, les Premières Nations, les Inuits et les Métis doivent composer avec des iniquités systémiques et persistantes en santé, qui sont associées à un héritage de colonialisme et de traumatisme intergénérationnel (ASPC, 2019). Ils présentent des taux de maladies infectieuses nettement plus élevés que les populations non autochtones, ce qui, par le fait même, accroît les risques de maladies infectieuses liées aux changements climatiques (Adekoya et coll., 2015). Si une plus grande sensibilité aux maladies infectieuses, qui peut être due à des facteurs génétiques ou à des taux élevés de comorbidités chroniques (Badawi et coll., 2018), a été observée (p. ex., sensibilité génétique à la grippe H1N1), les iniquités existantes en santé, telles que la disponibilité et l’accessibilité limitées de services de soins de santé acceptables, les logements surpeuplés et le manque d’information culturellement pertinente sur la prévention des maladies (Centre national de collaboration de la santé autochtone, 2016), peuvent aggraver la vulnérabilité (voir le chapitre 9 : Changements climatiques et équité en santé). En outre, le risque d’exposition pourrait aussi être plus élevé pour les Premières Nations, les Inuits et les Métis en raison de leur forte dépendance sur les aliments traditionnels et les activités axées sur la terre telles que la chasse. Par exemple, des éclosions de trichinose ont été associées à la consommation de mammifères marins dans les régions arctiques (Yansouni et coll., 2016) et l’augmentation de l’abondance de moustiques, de tiques et d’autres insectes piqueurs qui peuvent transmettre de nouvelles maladies à transmission vectorielle est de plus en plus préoccupante (voir le chapitre 2 : Changements climatiques et santé des Autochtones du Canada). L’emplacement géographique peut également accroître l’exposition, par exemple, le risque d’exposition à la rage du renard arctique est plus élevé pour les collectivités de l’Arctique.
Les politiques visant à traiter ces causes sous-jacentes de vulnérabilité ne relèvent généralement pas du mandat des organismes de santé publique (Adler et Newman, 2002). Toutefois, il est de plus en plus reconnu dans le domaine de la santé publique que les changements climatiques exacerberont les iniquités en santé (Friel, 2019). Les efforts visant à renforcer les déterminants de la santé devraient être combinés aux activités liées aux changements climatiques et à la santé (p. ex., les mesures d’adaptation) (voir le chapitre 9 : Changements climatiques et équité en santé).
6.4.6
Lacunes en matière d’adaptation et possibilités
Partager
6.4.6.1 Lacunes en matière d’adaptation
L’établissement des priorités en matière d’options d’adaptation pour gérer les risques de maladies infectieuses liés aux changements climatiques au Canada devrait tenir compte de multiples critères, y compris : le niveau des risques et leur caractère immédiat; la viabilité technique des options; les ressources humaines et financières; la compatibilité avec les politiques actuelles; et d’autres contraintes (Ebi et coll., 2006a; Ebi et coll., 2006b). Pour bon nombre des risques de maladies infectieuses associés aux changements climatiques dont il est question dans le présent chapitre, la surveillance, le suivi et le contrôle des maladies ne seront efficaces que si des mesures locales sont prises. Il sera donc important de permettre aux systèmes locaux de santé publique et de soins de santé d’entreprendre ces activités pour accroître la résilience face à ces menaces. Un examen de la planification nationale de l’adaptation aux impacts des changements climatiques sur les maladies infectieuses dans 14 pays membres de l’Organisation de coopération et de développement économiques a révélé un certain nombre de limites à la planification actuelle. Ces limites comprenaient la prise en compte négligeable des besoins des groupes de population qui subissent des impacts disproportionnés, la participation limitée des administrations locales à l’évaluation des risques pour la santé et l’insuffisance de la logistique et du soutien liés au financement, aux échéanciers pour les évaluations et à l’élaboration de plans d’adaptation (Panic et Ford, 2013). Le rapport souligne que ces limites doivent être prises en compte dans les plans d’adaptation canadiens. L’évaluation post-pandémique des réponses de la santé publique canadienne face à la pandémie de COVID-19 sera probablement très instructive en ce qui concerne les lacunes actuelles en matière de capacité d’adaptation.
De plus, l’adaptation aux maladies infectieuses liées au climat est entravée par les lacunes actuelles en matière de recherche. Pour de nombreuses maladies infectieuses liées au climat, il y a un manque de connaissance de leur écologie, peu de données de surveillance pour l’étalonnage des modèles de niche statistique/écologique et pour la validation des modèles prédictifs, et une absence de surveillance à long terme qui est nécessaire pour attribuer les changements du risque lié à la maladie aux changements climatiques (Ebi et coll., 2017). Il est particulièrement difficile de lutter efficacement contre les maladies transmises par les tiques dans l’environnement, et des recherches s’imposent dans ce domaine. Les examens de la portée et les examens systématiques concernant les risques liés à des maladies à transmission vectorielle particulières établies comme étant prioritaires par les méthodes d’ADMC permettent de cerner les lacunes en matière de connaissances et de recherche (Otten et coll., 2019).
La capacité de lutter contre les vecteurs diminue à l’échelle mondiale en raison de la résistance accrue aux insecticides, ce qui représentera un défi important à relever pour protéger la santé de la population canadienne. Cependant, des techniques telles que l’introduction de moustiques génétiquement modifiés qui contrôlent les populations en produisant des rejetons non viables et de moustiques porteurs de la bactérie Wolbachia dans leurs intestins (les rendant ainsi incompétents en tant que vecteurs), font l’objet de recherches. Ces techniques pourraient être mises à la disposition des responsables de la santé publique dans un proche avenir.
6.4.6.2 Possibilités d’adaptation
Des renseignements existants sur l’écologie et l’épidémiologie des maladies à transmission vectorielle qui menacent la population canadienne (Corrin et coll., 2018), de même que sur les méthodes de prévention et de contrôle et leur efficacité (Hierlihy et coll., 2019) sont fournis aux responsables de la santé publique de tous les ordres de gouvernement afin de les aider à élaborer des plans d’intervention. L’augmentation prévue du comportement épidémique des maladies transmises par les moustiques, telles que le VNO et le VEEE, signifie que les prévisions du risque de maladie fondées sur les prévisions météorologiques recommandées par le GIEC seraient utiles pour les systèmes d’alerte précoce. Beaucoup de progrès ont été réalisés pour ce qui est de l’élaboration de méthodes de prédiction des éclosions de VNO selon les prévisions météorologiques, mais la mise en œuvre des programmes de santé publique du Canada nécessitera d’autres travaux (Ogden et coll., 2019).
La possibilité croissante de transmission autochtone de maladies exotiques représente un défi pour les activités de santé publique et le système de santé canadien. En raison de l’absence de maladies exotiques au Canada, les professionnels de la santé ne les envisageaient pas comme des diagnostics possibles chez des patients qui n’avaient pas d’antécédents de voyage dans des régions endémiques bien connues. Un renforcement de la modélisation des risques de maladie améliorera la préparation aux événements d’émergence de maladies, et une surveillance accrue des nouvelles maladies infectieuses non endémiques chez les patients qui demandent des soins serait prudente.
De même, une surveillance renforcée est nécessaire pour les maladies à transmission vectorielle qui sont actuellement exotiques en Amérique du Nord, mais qui risquent d’être transportées au Canada par des voyageurs; celles qui sont absentes du Canada, mais qui risquent de s’étendre vers le nord en provenance des États-Unis; et celles qui sont endémiques au Canada, mais qui peuvent réapparaître sous forme d’éclosions. L’éducation des fournisseurs de soins de santé au sujet de l’émergence possible de ces nouvelles maladies au Canada sera importante tant pour les personnes touchées, qui pourront bénéficier d’un diagnostic et d’un traitement précoces, que pour la détection précoce des éclosions par la santé publique. Les approches de surveillance citoyenne, comme le programme eTick (eTick, 2019), sont également une occasion de plus en plus reconnue. Ces approches pourraient également être améliorées en intégrant le savoir et l’observation autochtones (Tomaselli et coll., 2017; Henri et coll., 2018).
Il est possible d’utiliser des « mégadonnées » et des technologies telles que l’intelligence artificielle (IA) qui aident à analyser les mégadonnées pour être en mesure de composer avec les risques croissants pour la santé liés aux maladies infectieuses. La génomique moderne est la première source de mégadonnées qui à être utilisée dans cette optique. La surveillance des maladies entériques intègre maintenant le séquençage du génome entier (SGE) grâce au réseau PulseNet au Canada, ainsi qu’aux États-Unis et en Europe. PulseNet permet une détection plus sensible des grappes de cas et des éclosions, et une plus grande capacité pour attribuer de façon fiable les sources d’infection (Gilmour et coll., 2013). La quantité considérable de données générées par le SGE signifie que des méthodes d’analyse bioinformatique complexes ont dû être élaborées pour l’analyse rapide des données (IRIDA, 2019). L’application du SGE à d’autres logiciels de diagnostic et de surveillance des maladies infectieuses, de même que le développement plus poussé de l’épidémiologie moléculaire en vue d’utiliser pleinement les données, se poursuivront à l’avenir et appuieront des efforts plus robustes pour s’attaquer à ces risques. Les méthodes de métagénomique et de métacodage à barres, qui permettent de cerner l’éventail des espèces présentes dans les échantillons complexes d’espèces mixtes, se prêtent à une surveillance élargie des moustiques et des maladies transmises par les moustiques, y compris les moustiques et les agents pathogènes exotiques. Grâce à ces méthodes, il est possible d’identifier un éventail des espèces de moustiques et des agents pathogènes transmis par les moustiques capturés dans les pièges à moustiques pendant la surveillance des moustiques, plutôt que simplement le VNO. Ces méthodes pourraient également révolutionner la détection d’agents pathogènes dans d’autres échantillons d’ADN environnemental provenant de l’eau potable, de l’eau utilisée à des fins récréatives et du sol.
La deuxième source de mégadonnées provient des médias Web et des médias sociaux; ces données sont obtenues au moyen de méthodes de recherche sur le Web (Mukhi et coll., 2016). Cette source peut fournir des signaux précoces d’événements d’émergence de maladies afin d’améliorer les efforts de gestion d’organisations comme le RMISP. La troisième source de mégadonnées sont les données d’observation de la Terre obtenues par satellite. Elles fournissent des approximations pour les données environnementales comme les conditions météorologiques, le climat, les habitats et l’aménagement du territoire, qui peuvent être utilisées avec la modélisation des risques pour déterminer où les risques d’émergence et de réémergence de maladies sensibles aux facteurs environnementaux peuvent se produire (Ceccato et coll., 2018). Il existe de nombreux exemples de données d’observation de la Terre utilisées pour déterminer le risque de maladie (en particulier la maladie de Lyme) au Canada sous forme de cartes statiques des risques (figure 6.9). Les données d’observation de la Terre deviennent de plus en plus « volumineuses » en raison de l’augmentation du nombre et des types de satellites, la résolution spatiale des données étant de plus en plus précise. Dans un proche avenir, une plus grande fréquence d’images pourrait permettre de déterminer en temps réel les risques de maladies à transmission vectorielle comme le VNO. Du moins en théorie, les données d’observation de la Terre, synthétisées par modélisation des risques afin de produire de l’information sur les risques, pourraient remplacer la surveillance comme moyen de déclencher des interventions de santé publique bien avant que les cas de maladie n’apparaissent chez l’humain (Ogden et coll., 2019).
Cartes des risques liés à la maladie de Lyme dans l’Est du Canada en fonction du risque de populations d’Ixodes scapularis, grâce aux données d’observation de la Terre sur la qualité du climat et de l’habitat.
Figure 6.9
Ces cartes des risques mesurent l’adéquation environnementale pour les tiques vectrices de la maladie de Lyme Ixodes scapularis dans l’est et le centre du Canada à l’aide de données d’observation de la Terre approximatives pour la température (degrés-jours cumulatifs annuels supérieurs à 0 °C) et l’habitat forestier. L’adéquation environnementale est nulle là où il n’y a pas de terrain boisé (zones grises plus claires) et là où le climat est trop froid (zones bleu foncé). Dans les zones où se trouve un terrain boisé et où le climat est suffisamment chaud, le risque d’occurrence des populations de tiques varie de zones à faible risque (zones bleu pâle) à des zones à risque modéré, modéré-élevé et élevé (zones vertes, orange et rouges, respectivement). L’échelle d’adéquation pour la tique selon les conditions de température (avec des valeurs de 0 à 1 500) est obtenue à partir d’un modèle mathématique des populations d’I. scapularis. Les données d’observation de la Terre servant à établir cette carte n’étaient pas disponibles pour certaines parties du Nunavut et des Territoires du Nord-Ouest (zones gris foncé).
Source
Kotchi et coll., 2019.
Beaucoup de travail a été fait pour déterminer et quantifier les effets des conditions météorologiques sur les moustiques vecteurs du VNO et sur le risque de transmission du VNO au Canada (voir la section 6.3.1.3 Maladies endémiques transmises par les moustiques au Canada), ce qui servirait de base à l’élaboration de prévisions sur les éclosions de VNO. La combinaison des données météorologiques et des données d’observation de la Terre sur l’habitat dans les modèles prédictifs des systèmes d’information géographique permettrait de faire des prévisions à haute résolution spatiotemporelle.
Bon nombre des défis posés par les nouveaux vecteurs et nouvelles maladies qui menacent la santé publique dans le contexte des changements climatiques sont communs à d’autres disciplines telles que l’agriculture (y compris la santé végétale et animale), la gestion de l’environnement et de la biodiversité, et les ressources naturelles qui devraient subir des invasions d’espèces (infectieuses ou autres) en raison des changements climatiques, entre autres facteurs. La collaboration en matière d’activités d’évaluation, de surveillance et de gestion des risques au moyen d’une approche « Un monde, une santé » (ou « Une seule santé ») robuste peut présenter des avantages importants par rapport à la gestion cloisonnée des risques propres au secteur (Ogden et coll., 2019). Cela nécessitera une collaboration plus systématique entre les disciplines et les ministères, qui sont tous confrontés à l’émergence de menaces pour la santé (humaine, animale et environnementale) associées aux changements climatiques.
6.5
Lacunes sur le plan des connaissances
Partager
Les lacunes actuelles au chapitre des connaissances liées aux impacts des changements climatiques sur les maladies infectieuses au Canada nuisent à l’élaboration d’évaluations locales et régionales des changements climatiques et de la vulnérabilité en matière de santé, de même que de plans d’adaptation. Elles empêchent également les responsables de la santé publique de détecter l’émergence et la réémergence de maladies infectieuses, ainsi que de prévenir et de contrôler une telle émergence. Les principales lacunes sont décrites ci-dessous.
Il faut mieux connaître l’écologie et l’épidémiologie des maladies infectieuses et des arthropodes vecteurs pour appuyer l’élaboration d’évaluations des risques et de systèmes d’alerte précoce. En outre, une surveillance systématique des cas humains, des arthropodes vecteurs, des animaux sentinelles infectés ou des échantillons environnementaux, ou le recours à des méthodes de science citoyenne, à des échelles spatiotemporelles utiles, est nécessaire pour cerner les maladies et les vecteurs émergents. Bien que cela se produise déjà dans une certaine mesure, des méthodes de surveillance (p. ex., méthodes de métagénomique) qui détectent de multiples agents pathogènes et vecteurs possiblement émergents sont nécessaires pour passer de la surveillance ciblée d’une maladie unique à des programmes qui s’attaquent à une vaste gamme d’agents pathogènes et de vecteurs qui menacent la population canadienne. Il faut mettre au point des méthodes et des algorithmes de diagnostic des maladies infectieuses afin que les maladies actuellement considérées comme étant exotiques au Canada soient détectées plus facilement chez les Canadiens et les Canadiennes qui n’ont pas voyagé dans des pays où ces maladies sont connues depuis longtemps. Une surveillance à long terme sera nécessaire pour appuyer l’attribution aux effets des changements climatiques.
Il existe un large éventail de maladies infectieuses dont la sensibilité aux conditions météorologiques et climatiques est connue, mais pour bon nombre d’entre elles, l’évaluation des impacts des changements climatiques n’a pas été effectuée. Au Canada, les évaluations qui ont été effectuées ont principalement porté sur les maladies à transmission vectorielle. Les méthodes utilisées pour les maladies à transmission vectorielle doivent être appliquées à un plus large éventail de maladies infectieuses, y compris les maladies transmises de personne à personne, et les maladies infectieuses provenant de sources environnementales qui sont transmises par le vent et l’air.
Pour attribuer l’émergence et la réémergence de maladies infectieuses au Canada aux changements climatiques, il faut disposer de renseignements qui permettent d’analyser la force statistique de l’association, la cohérence entre les études, la spécificité des effets du climat, une concordance appropriée dans le temps des changements climatiques et de l’émergence des maladies, le gradient biologique (des changements climatiques plus importants donnent lieu à des effets plus marqués) et la plausibilité (Hill, 1965).
Il faut resserrer les liens entre les efforts de santé publique pour comprendre les maladies infectieuses et chroniques et y réagir. On reconnaît de plus en plus que les maladies infectieuses peuvent entraîner des maladies chroniques (O’Connor et coll., 2006) et que les maladies infectieuses sont plus graves chez les personnes atteintes de maladies chroniques (Badawi et coll., 2018). De plus, le statut socioéconomique a une incidence sur la vulnérabilité aux maladies infectieuses et chroniques, de même que sur la perception du risque, la connaissance des mesures de protection et la volonté de les utiliser (Bouchard et coll., 2018). L’évolution démographique selon l’âge au Canada, comme dans d’autres pays à revenu élevé, associée à l’incidence croissante des maladies chroniques dans ces populations (Dye, 2014), et les effets prévus des changements climatiques sur les maladies infectieuses et chroniques signifient que les responsables de la santé publique et les chercheurs doivent comprendre les impacts des changements climatiques sur ces maladies, les prendre en compte et y réagir de manière coordonnée.
Il existe des lacunes considérables dans la capacité du Canada à prévenir et à contrôler les maladies à transmission vectorielle. Ces lacunes comprennent une connaissance limitée d’un éventail de mesures efficaces pour contrôler les vecteurs dans l’environnement et pour permettre aux gens de se protéger, de même qu’un manque de protocoles établis pour les programmes de lutte contre les maladies à transmission vectorielle destinés aux intervenants de première ligne en santé publique. La nécessité d’innover en matière de prévention et de contrôle est mise en évidence par les défis croissants de la résistance aux antimicrobiens et de la résistance aux insecticides à l’échelle internationale.
Il faut valider les systèmes d’alerte précoce (Morin et coll., 2018) pour les maladies infectieuses, comme les maladies transmises par les moustiques, qui peuvent devenir plus épidémiques en raison des changements climatiques et qui menacent de se propager rapidement au Canada, comme ce fût le cas lors de l’introduction du VNO. En outre, les collectivités canadiennes aux niveaux municipal, provincial et territorial ont besoin d’évaluations exhaustives de la vulnérabilité aux effets des changements climatiques sur tous les aspects de la santé, comme les maladies et les décès liés à la chaleur, les maladies chroniques et les risques de maladies infectieuses. Ces évaluations constituent la première étape en vue de réduire les risques futurs de maladie, ainsi que de cerner les lacunes dans les connaissances et le système de santé publique, qui doivent être comblées par des investissements ou de la recherche (voir le chapitre 10 : Adaptation et résilience des systèmes de santé).
Pour combler les lacunes existantes en matière de connaissances à l’appui d’une adaptation efficace, il faut une collaboration directe et bien planifiée avec la santé publique et les utilisateurs finaux (Nyström et coll., 2018). Les initiatives de collaboration, comme celle établie entre l’ASPC et l’Université de Montréal dans leur Groupe de recherche en épidémiologie des zoonoses et santé publique, accéléreraient la capacité d’innover et de réagir aux menaces émergentes liées aux maladies (GREZOSP, 2019). Toutefois, de telles collaborations nécessitent des investissements à long terme, l’harmonisation des buts et des objectifs entre les partenaires, des compétences mutuelles, le respect et la confiance, ainsi qu’une proximité géographique pour réussir (Rycroft-Malone et coll., 2015).
6.6
Conclusion
Partager
Depuis la publication en 2008 de la première évaluation canadienne exhaustive de la vulnérabilité en matière de santé et de l’adaptation aux changements climatiques, des efforts considérables ont été déployés pour évaluer les risques liés aux maladies émergentes (en particulier les maladies à transmission vectorielle), mettre en œuvre la surveillance et diffuser de l’information sur la prévention et le contrôle efficaces aux professionnels de la santé publique et au membres du grand public. Depuis, le vecteur de la maladie de Lyme s’est propagé au Canada comme prévu et a entraîné l’émergence rapide de la maladie de Lyme et d’autres maladies transmises par les tiques. Le VEEE s’est également répandu au Canada, une éclosion de VNO s’est produite dans l’est du Canada et l’Ae. albopictus s’est établi dans le sud de l’Ontario. Ces événements confirment le risque accru de propagation vers le nord de vecteurs et de maladies à transmission vectorielle en provenance des États-Unis, et d’épidémies de maladies endémiques à transmission vectorielle causées par les conditions météorologiques, tel qu’il est mentionné dans l’évaluation de 2008. Ils mettent également en lumière la réalité de l’émergence et de la réémergence des maladies à transmission vectorielle découlant des changements climatiques.
L’apparition de la première infection par le virus chikungunya, puis par le virus Zika, dans les Amériques, a également accru les préoccupations des responsables de la santé publique au Canada et à l’étranger au sujet des risques associés à la propagation de maladies exotiques venant de loin, à leur introduction et à leur transmission endémique. La question de savoir si ces événements d’émergence de maladies avaient été causés ou non par les changements climatiques, en tout ou en partie, a fait l’objet d’un débat (Paz et Semenza, 2016). D’une façon ou d’une autre, l’identification dans le sud de l’Ontario de populations d’Ae. albopictus, un vecteur de ces deux maladies, signifie que la possibilité de transmission autochtone de maladies exotiques au Canada ne relève plus d’une hypothèse. Les données présentées dans le présent chapitre indiquent que la dengue et la fièvre de la vallée du Rift sont les maladies exotiques à transmission vectorielle les plus graves qui pourraient apparaître au Canada suite à une dispersion sur de longues distances. De plus, bien que l’évaluation de 2008 ait fait état de maladies sensibles au climat, il n’y a pas eu d’événements majeurs d’émergence ou de réémergence associés à la peste et aux zoonoses transmises directement comme le hantavirus depuis cette époque. Toutefois, aucun effort n’a été déployé pour explorer plus à fond les effets futurs possibles des changements climatiques sur ces agents pathogènes.
Les changements climatiques et les conditions météorologiques peuvent accroître les risques liés aux maladies infectieuses transmises d’une personne à l’autre et de maladies infectieuses transmises par l’air provenant de sources environnementales. De façon générale, des éclosions ou des événements d’émergence de ces maladies sensibles au climat, comme la paralysie flasque aiguë et la blastomycose, se sont produits au Canada ou en Amérique du Nord (Trudel et coll., 2014; Elrick et coll., 2019). Par conséquent, il est justifié de mener davantage de recherches sur les effets possibles des changements climatiques sur ces maladies.
À l’instar d’autres pays à revenu élevé, la capacité d’adaptation au Canada pour gérer les risques de maladies infectieuses émergentes et réémergentes est considérée comme étant excellente. Cela est attribuable à la contribution combinée des systèmes de santé publique nationaux et internationaux, de même qu’à la robustesse des services de soins de santé et à la capacité technologique. Néanmoins, des lacunes subsistent au chapitre des connaissances et des activités de santé publique. Celles-ci incluent la nécessité de réaliser des évaluations supplémentaires des effets des changements climatiques sur les risques liés aux maladies infectieuses; d’effectuer des évaluations plus systématiques de la vulnérabilité par les municipalités, les provinces et les territoires; d’améliorer la surveillance, y compris les programmes de surveillance à long terme des effets des changements climatiques; d’accroître la capacité de contrôle. Ces lacunes ne peuvent être comblées que par un accroissement de la recherche et des investissements dans les interventions en santé publique, une collaboration plus directe des utilisateurs finaux de la santé publique et des chercheurs universitaires, et l’application d’approches multidisciplinaires de type « Un monde, une santé » (ou « Une seule santé »).
Footnotes
Les aliments traditionnels inuits, aussi appelés « aliments prélevés dans la nature », font partie intégrante de l’identité et de la culture inuites, sont une source importante de nutriments et contribuent à la santé et au bien-être des personnes et des collectivités. Ils comprennent les animaux marins (p. ex., morses, phoques), les caribous, les oiseaux, les poissons et les aliments fourragers.
↩
References
Acheson, E. S., Galanis, E., Bartlett, K., Klinkenberg, B. (2019). Climate classification system-based determination of temperate climate detection of Cryptococcus gattii sensu lato. Emerging Infectious Diseases, 25(9), 1723-1726.
↩
Acheson, E. S., Galanis, E., Bartlett, K., Mak, S., et Klinkenberg, B. (2018). Searching for clues for eighteen years: Deciphering the ecological determinants of Cryptococcus gattii on Vancouver Island, British Columbia. Medical Mycology, 56(2), 129-144. https://doi.org/10.1093/mmy/myx037 ↩
Adekoya N, Truman B, Landen M; Centers for Disease Control and Prevention. (2015). Incidence of notifiable diseases among American Indians/Alaska Natives – United States, 2007-2011. MMWR Morb Mortal Weekly Report, 64(1), 16-9.
↩
Adler NE, Newman K. (2002). Socioeconomic disparities in health: pathways and policies. Health Affairs (Project Hope), 21(2), 60-76. https://doi.org/10.1377/hlthaff.21.2.60 ↩
Aenishaenslin, C., Simon, A., Forde, T., Ravel, A., Proulx, J.-F., Fehlner-Gardiner, C., . . . Belanger, D. (2014). Characterizing rabies epidemiology in remote Inuit communities in Québec, Canada: A « One Health » approach. EcoHealth, 11(3), 343-355. https://doi.org/10.1007/s10393-014-0923-1 ↩
Alarcon Falconi, T. M., Cruz, M. S., et Naumova, E. N. (2018). The shift in seasonality of legionellosis in the USA. Epidemiology et Infection, 146(14), 1824-1833. https://doi.org/10.1017/S0950268818002182 ↩
Altizer, S., Ostfeld, R. S., Johnson, P. T. J., Kutz, S., et Harvell, C. D. (2013). Climate change and infectious diseases: From evidence to a predictive framework. Science, 341(6145), 514-519. https://doi.org/10.1126/science.1239401 ↩
Ampofo, K., Gesteland, P. H., Bender, J., Mills, M., Daly, J., Samore, M., . . . Srivastava, R. (2006). Epidemiology, complications, and cost of hospitalization in children with laboratory-confirmed influenza infection. Pediatrics, 118(6), 2409-2417. https://doi.org/10.1542/peds.2006-1475 ↩
Andersen, K. G., Rambaut, A., Lipkin, W. I., Holmes, E. C., et Garry, R. F. (2020). The proximal origin of SARS-CoV-2. Nature medicine, 26(4), 450–452. https://doi.org/10.1038/s41591-020-0820-9 ↩
Anderson, H., Honish, L., Taylor, G., Johnson, M., Tovstiuk, C., Fanning, A., Tyrrell, G., Rennie, R., Jaipaul, J., Sand, C., et Probert, S. (2006). Histoplasmosis cluster, golf course, Canada. Emerging infectious diseases, 12(1), 163–165. https://doi.org/10.3201/eid1201.051083 ↩
Anderson, R. M., et May, R. M. (1991). Infectious diseases of humans: Dynamics and control. Oxford, England: Oxford University Press.
↩
Andreadis, T. G., Anderson, J. F., Armstrong, P. M., et Main, A. J. (2008). Isolations of Jamestown Canyon virus (Bunyaviridae: Orthobunyavirus) from field-collected mosquitoes (Diptera: Culicidae) in Connecticut, USA: A ten-year analysis, 1997–2006. Vector-Borne and Zoonotic Diseases, 8(2), 175-188. https://doi.org/10.1089/vbz.2007.0169 ↩
Anyamba, A., Linthicum, K. J., Small, J. L., Collins, K. M., Tucker, C. J., Pak, E. W., . . . Russell, K. L. (2012). Climate teleconnections and recent patterns of human and animal disease outbreaks. PLOS Neglected Tropical Diseases, 6(1), 1-14. https://doi.org/10.1371/journal.pntd.0001465 ↩
Arksey, H., et O’Malley, L. (2005). Scoping studies: Towards a methodological framework. International Journal of Social Research Methodology, 8(1), 19-32. https://doi.org/10.1080/1364557032000119616 ↩
Armstrong, P. M., et Andreadis, T. G. (2010). Eastern equine encephalitis virus in mosquitoes and their role as bridge vectors. Emerging Infectious Diseases, 16(12), 1869-1874. https://doi.org/10.3201/eid1612.100640 ↩
Artsob, H. (1988). Powassan encephalitis. In T. P. Monath (Ed.), The arboviruses: Epidemiology and ecology (Vol. 4, pp. 29-49). Boca Raton, FL: CRC Press.
↩
Artsob, H. (2000). Arthropod-borne disease in Canada: A clinician’s perspective from the ‘Cold Zone’. Paediatrics et Child Health, 5(4), 206-212. https://doi.org/10.1093/pch/5.4.206 ↩
Artsob, H., Gubler, D. J., Enria, D. A., Morales, M. A., Pupo, M., Bunning, M. L., et Dudley, J. P. (2009). West Nile virus in the New World: Trends in the spread and proliferation of West Nile virus in the Western Hemisphere. Zoonoses and Public Health, 56(6-7), 357-369. https://doi.org/10.1111/j.1863-2378.2008.01207.x ↩
Asad, H., et Carpenter, D. O. (2018). Effects of climate change on the spread of Zika virus: A public health threat. Reviews on Environmental Health, 33(1), 31-42. https://doi.org/10.1515/reveh-2017-0042 ↩
Ashraf, U., Ye, J., Ruan, X., Wan, S., Zhu, B., et Cao, S. (2015). Usutu virus: An emerging flavivirus in Europe. Viruses, 7(1), 219-238. https://doi.org/10.3390/v7010219 ↩
Aune, K., Rhyan, J. C., Russell, R., Roffe, T. J., et Corso, B. (2012). Environmental persistence of Brucella abortus in the Greater Yellowstone Area. The Journal of Wildlife Management, 76(2), 253-261. https://doi.org/10.1002/jwmg.274 ↩
Ayers, V. B., Huang, Y.-J. S., Lyons, A. C., Park, S. L., Higgs, S., Dunlop, J. I., . . . Vanlandingham, D. L. (2018). Culex tarsalis is a competent vector species for Cache Valley virus. Parasites et Vectors, 11, 1-6. https://doi.org/10.1186/s13071-018-3103-2 ↩
Azam, D., Ukpai, O. M., Said, A., Abd-Allah, G. A., et Morgan, E. R. (2012). Temperature and the development and survival of infective Toxocara canis larvae. Parasitology Research, 110(2), 649-656. https://doi.org/10.1007/s00436-011-2536-8 ↩
Azar, M. M., Assi, R., Relich, R. F., Schmitt, B. H., Norris, S., Wheat, L. J., et Hage, C. A. (2015). Blastomycosis in Indiana. Chest, 148(5), 1276-1284. https://doi.org/10.1378/chest.15-0289 ↩
Badawi, A., Velummailum, R., Ryoo, S. G., Senthinathan, A., Yaghoubi, S., Vasileva, D., . . . Arora, P. (2018). Prevalence of chronic comorbidities in dengue fever and West Nile virus: A systematic review and meta-analysis. PLOS ONE, 13(7), 1-27. https://doi.org/10.1371/journal.pone.0200200 ↩
Baumgardner, D. J., et Burdick, J. S. (1991). An outbreak of human and canine blastomycosis. Reviews of Infectious Diseases, 13(5), 898-905. https://doi.org/10.1093/clinids/13.5.898 ↩
Baumgardner, D. J., Paretsky, D. P., Baeseman, Z. J., et Schreiber, A. (2011). Effects of season and weather on blastomycosis in dogs: Northern Wisconsin, USA. Medical Mycology, 49(1), 49-55. https://doi.org/10.3109/13693786.2010.488658 ↩
Beard, C. B., Eisen, R. J., Barker, C. M., Garofalo, J. F., Hahn, M., Hayden, M., . . . Schramm, P. J. (2016). Vector-borne diseases. In A. Crimmins, J. Balbus, J. L. Gamble, C. B. Beard, J. E. Bell, D. Dodgen, R. J. Eisen, N. Fann, M. D. Hawkins, S. C. Herring, L. Jantarasami, D. M. Mills, S. Saha, M. C. Sarofim, J. Trtanj, et L. Ziska (Eds.), The impacts of climate change on human health in the United States: A scientific assessment (pp. 129-156). Washington, DC: U.S. Global Change Research Program. https://doi.org/10.7930/J0765C7V ↩
Beaudoin, M., et Gosselin, P. (2016). An effective public health program to reduce urban heat islands in Québec, Canada. Pan American Journal of Public Health, 40(3), 160-166. Consulté sur le site : http://iris.paho.org/xmlui/handle/123456789/31230 ↩
Beauté, J., Sandin, S., Uldum, S. A., Rota, M. C., Brandsema, P., Giesecke, J., et Sparén, P. (2016). Short-term effects of atmospheric pressure, temperature, and rainfall on notification rate of community-acquired Legionnaires’ disease in four European countries. Epidemiology et Infection, 144(16), 3483-3493. https://doi.org/10.1017/S0950268816001874 ↩
Ben Ari, T., Gershunov, A., Gage, K. L., Snäll, T., Ettestad, P., Kausrud, K. L., et Stenseth, N. C. (2008). Human plague in the USA: The importance of regional and local climate. Biology Letters, 4(6), 737-740. https://doi.org/10.1098/rsbl.2008.0363 ↩
Ben Ari, T., Gershunov, A., Tristan, R., Cazelles, B., Gage, K., et Stenseth, N. C. (2010). Interannual variability of human plague occurrence in the western United States explained by tropical and North Pacific Ocean climate variability. The American Journal of Tropical Medicine and Hygiene, 83(3), 624-632. https://doi.org/10.4269/ajtmh.2010.09-0775 ↩
Bern, C., et Montgomery, S. P. (2009). An estimate of the burden of Chagas disease in the United States. Clinical Infectious Diseases, 49(5), e52-e54. https://doi.org/10.1086/605091 ↩
Berrang-Ford, L., Maclean, J. D., Gyorkos, T. W., Ford, J. D., et Ogden, N. H. (2009). Climate change and malaria in Canada: A systems approach. Interdisciplinary Perspectives on Infectious Diseases, 2009, 1-13. https://doi.org/10.1155/2009/385487 ↩
Berry, P. (2008). Vulnérabilités, adaptation et capacité d’adaptation au Canada. Dans J. Séguin (éd.), Santé et changements climatiques: évaluation des vulnérabilitiés et de la capacité d’adaptation au Canada (pages 367 à 448). Ottawa, ON: Gouvernement of Canada. Consulté sur le site https://publications.gc.ca/site/fra/9.635906/publication.html ↩
Berry, P., Clarke, K.-L., Fleury, M. D., et Parker, S. (2014a). Santé humaine. Dans F. J. Warren et D. S. Lemmen (éd.), Vivre avec les changements climatiques au Canada : perspectives des secteurs relatives aux impacts et à l’adaptation (pages 191 à 232). Ottawa, ON: Gouvernement du Canada. Consulté sur le site https://publications.gc.ca/site/fra/9.643395/publication.html ↩
Berry, P., Patterson, J., Buse, C. (2014b). Assessment of Vulnerability to the Health Impacts of Climate Change in Middlesex-London. Consulté sur le site : https://www.healthunit.com/climate-change ↩
Berteaux, D., de Blois, S., Angers, J.-F., Bonin, J., Casajus, N., Darveau, M., . . . Vescovi, L. (2010). The CC-Bio project: Studying the effects of climate change on Quebec biodiversity. Diversity, 2(11), 1181-1204. https://doi.org/10.3390/d2111181 ↩
Berteaux, D., et Stenseth, N. C. (2006). Measuring, understanding and projecting the effects of large-scale climatic variability on mammals. Climate Research, 32(2), 95-97. Consulté sur le site : https://www.int-res.com/articles/cr_oa/c032p095.pdf ↩
Biggs, H. M., Behravesh, C. B., Bradley, K. K., Dahlgren, F. S., Drexler, N. A., Dumler, J. S., . . . Traeger, M. S. (2016). Diagnosis and management of tickborne rickettsial diseases: Rocky Mountain spotted fever and other spotted fever group Rickettsioses, Ehrlichioses, and Anaplasmosis – United States. MMWR Recommendations and Reports, 65(2), 1-44. https://doi.org/10.15585/mmwr.rr6502a1 ↩
Blancher, P., Cadman, M. D., Pond, B. A., Couturier, A. R., Dunn, E. H., Francis, C. M., et Rempel, R. S. (2008). Changements dans la répartition des oiseaux depuis le premier atlas. In M. D. Cadman, D. A. Sutherland, G. G. Beck, D. Lepage, et A. R. Couturier (éds.), Atlas des oiseaux nicheurs de l’Ontario, 2001-2005 (pp. 32-48). Toronto, Ontario: Études d’oiseaux Canada, Environnement Canada, Ontario Field Ornithologists, ministère des richesses naturelles de l’Ontario, et Ontario Nature.
↩
Bouchard, C., Aenishaenslin, C., Rees, E. E., Koffi, J. K., Pelcat, Y., Ripoche, M., . . . Leighton, P. A. (2018). Integrated social-behavioral and ecological risk maps to prioritize local public health responses to Lyme disease. Environmental Health Perspectives, 126(4). https://doi.org/10.1289/EHP1943 ↩
Bouchard, C., Dibernardo, A., Koffi, J., Wood, H., Leighton, P. A., et Lindsay, L. R. (2019). Increased risk of tick-borne diseases with climate and environmental changes. Canada Communicable Disease Report, 45(4), 83-89. https://doi.org/10.14745/ccdr.v45i04a02 ↩
Bouchard, C., Leonard, E., Koffi, J. K., Pelcat, Y., Peregrine, A., Chilton, N., . . . Ogden, N. H. (2015). The increasing risk of Lyme disease in Canada. The Canadian Veterinary Journal, 56(7), 693-699. Consulté sur le site : https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4466818/ ↩
Bouri, N., Sell, T. K., Franco, C., Adalja, A. A., Henderson, D. A., et Hynes, N. A. (2012). Return of epidemic dengue in the United States: Implications for the public health practitioner. Public Health Reports, 127(3), 259-266. https://doi.org/10.1177/003335491212700305 ↩
Brachman, P. S., et Abrutyn, E. (Eds.). (2009). Bacterial infections of humans: Epidemiology and control (4th ed.). New York, NY: Springer.
↩
Brady, O. J., Johansson, M. A., Guerra, C. A., Bhatt, S., Golding, N., Pigott, D. M., . . . Hay, S. I. (2013). Modelling adult Aedes aegypti and Aedes albopictus survival at different temperatures in laboratory and field settings. Parasites et Vectors, 6, 1-12. https://doi.org/10.1186/1756-3305-6-351 ↩
Brierley, L., Vonhof, M. J., Olival, K. J., Daszak, P., et Jones, K. E. (2016). Quantifying global drivers of zoonotic bat viruses: A process-based perspective. The American Naturalist, 187(2), E53-E64. https://doi.org/10.1086/684391 ↩
Brook, R. K., Kutz, S. J., Veitch, A. M., Popko, R. A., Elkin, B. T., et Guthrie, G. (2009). Fostering community-based wildlife health monitoring and research in the Canadian North. EcoHealth, 6(2), 266-278. https://doi.org/10.1007/s10393-009-0256-7 ↩
Brown, E. M., McTaggart, L. R., Dunn, D., Pszczolko, E., Tsui, K. G., Morris, S. K., . . . Richardson, S. E. (2018). Epidemiology and geographic distribution of blastomycosis, histoplasmosis, and coccidioidomycosis, Ontario, Canada, 1990–2015. Emerging Infectious Diseases, 24(7), 1257-1266. https://doi.org/10.3201/eid2407.172063 ↩
Brownstein, J. S., Holford, T. R., et Fish, D. (2005a). Effect of climate change on Lyme disease risk in North America. EcoHealth, 2(1), 38-46. https://doi.org/10.1007/s10393-004-0139-x ↩
Brownstein, J. S., Skelly, D. K., Holford, T. R., et Fish, D. (2005b). Forest fragmentation predicts local scale heterogeneity of Lyme disease risk. Oecologia, 146(3), 469-475. https://doi.org/10.1007/s00442-005-0251-9 ↩
Bullard, J. M. P., Ahsanuddin, A. N., Perry, A. M., Lindsay, L. R., Iranpour, M., Dibernardo, A., et Van Caeseele, P. G. (2014). The first case of locally acquired tick-borne Babesia microti infection in Canada. Canadian Journal of Infectious Diseases and Medical Microbiology, 25(6), e87-e89. https://doi.org/10.1155/2014/209521 ↩
Bush, E., et Lemmen D. (éd.). (2019). Rapport sur le climat changeant du Canada. Ottawa, ON: Gouvernement du Canada. Consulté sur le site https://changingclimate.ca/CCCR2019/fr/ ↩
Cable, J., Barber, I., Boag, B., Ellison, A. R., Morgan, E. R., Murray, K., . . . Booth, M. (2017). Global change, parasite transmission and disease control: Lessons from ecology. Philosophical Transactions of the Royal Society B: Biological Sciences, 372(1719), 1-17. https://doi.org/10.1098/rstb.2016.0088 ↩
Caini, S., Spreeuwenberg, P., Donker, G., Korevaar, J., et Paget, J. (2018). Climatic factors and long-term trends of influenza-like illness rates in the Netherlands, 1970–2016. Environmental Research, 167, 307-313. https://doi.org/10.1016/j.envres.2018.07.035 ↩
Campbell, L. P., Luther, C., Moo-Llanes, D., Ramsey, J. M., Danis-Lozano, R., et Peterson, A. T. (2015). Climate change influences on global distributions of dengue and chikungunya virus vectors. Philosophical Transactions of the Royal Society B: Biological Sciences, 370(1665), 1-9. https://doi.org/10.1098/rstb.2014.0135 ↩
Carlsen, B., et Glenton, C. (2016). The swine flu vaccine, public attitudes, and researcher interpretations: A systematic review of qualitative research. BMC Health Services Research, 16, 1-15. https://doi.org/10.1186/s12913-016-1466-7 ↩
Carmona-Castro, O., Moo-Llanes, D. A., et Ramsey, J. M. (2018). Impact of climate change on vector transmission of Trypanosoma cruzi (Chagas, 1909) in North America. Medical and Veterinary Entomology, 32(1), 84-101. https://doi.org/10.1111/mve.12269 ↩
Castillo, C. G., Kauffman, C. A., et Miceli, M. H. (2016). Blastomycosis. Infectious Disease Clinics of North America, 30(1), 247-264. https://doi.org/10.1016/j.idc.2015.10.002 ↩
Ceccato, P., Ramirez, B., Manyangadze, T., Gwakisa, P., et Thomson, M. C. (2018). Data and tools to integrate climate and environmental information into public health. Infectious Diseases of Poverty, 7, 1-11. https://doi.org/10.1186/s40249-018-0501-9 ↩
Centers for Disease Control and Prevention. (2018a). La Crosse encephalitis: Epidemiology et geographic distribution. Consulté sur le site : https://www.cdc.gov/lac/tech/epi.html ↩
Centre de collaboration nationale de la santé autochtone. (2016). Les déterminants de la prévalence et de la gravité de l’infection par le virus de la grippe au sein des populations autochtones du Canada. Prince George, Colombie-Britannique: Centre de collaboration nationale de la santé autochtone
↩
Centre for Food-borne, Environmental and Zoonotic Infectious Diseases. (2017). Synopsis: Lyme Disease in Canada – A Federal Framework. Canada communicable disease report, 43(10), 212-214. https://doi.org/10.14745/ccdr.v43i10a04 ↩
Chambers, D., Périé, C., Casajus, N., et de Blois, S. (2013). Challenges in modelling the abundance of 105 tree species in eastern North America using climate, edaphic, and topographic variables. Forest Ecology and Management, 291, 20-29. https://doi.org/10.1016/j.foreco.2012.10.046 ↩
Charron, D., Fleury, M., Lindsay, L. R., Ogden, N., et Schuster, C. J. (2008). Répercussions des changements climatiques sur les maladies transmises par l’eau, les aliments, les vecteurs et les rongeurs. Dans J. Séguin (éd.), Santé et changements climatiques: évaluation des vulnérabilitiés et de la capacité d’adaptation au Canada (pages 171 à 210). Ottawa, ON: Gouvernement of Canada. Consulté sur le site https://publications.gc.ca/site/fra/9.635906/publication.html ↩
Chattopadhyay, I., Kiciman, E., Elliott, J. W., Shaman, J. L., et Rzhetsky, A. (2018). Conjunction of factors triggering waves of seasonal influenza. eLife, 7, 1-44. https://doi.org/10.7554/eLife.30756 ↩
Chen, C. C., Jenkins, E., Epp, T., Waldner, C., Curry, P. S., et Soos, C. (2013). Climate change and West Nile virus in a highly endemic region of North America. International Journal of Environmental Research and Public Health, 10(7), 3052-3071. https://doi.org/10.3390/ijerph10073052 ↩
Chen, N.-T., Chen, M.-J., Guo, C.-Y., Chen, K.-T., et Su, H.-J. (2014a). Precipitation increases the occurrence of sporadic Legionnaires’ disease in Taiwan. PLOS ONE, 9(12), 1-11. https://doi.org/10.1371/journal.pone.0114337 ↩
Chen, S. C.-A., Meyer, W., et Sorrell, T. C. (2014b). Cryptococcus gattii infections. Clinical Microbiology Reviews, 27(4), 980-1024. https://doi.org/10.1128/CMR.00126-13 ↩
Cheng, J., Wu, J., Xu, Z., Zhu, R., Wang, X., Li, K., . . . Su, H. (2014). Associations between extreme precipitation and childhood hand, foot and mouth disease in urban and rural areas in Hefei, China. Science of the Total Environment, 497-498, 484-490. https://doi.org/10.1016/j.scitotenv.2014.08.006 ↩
Choi, K. M., Christakos, G., et Wilson, M. L. (2006). El Niño effects on influenza mortality risks in the state of California. Public Health, 120(6), 505-516. https://doi.org/10.1016/j.puhe.2006.01.011 ↩
Chretien, J.-P., Anyamba, A., Bedno, S. A., Breiman, R. F., Sang, R., Sergon, K., . . . Linthicum, K. J. (2007). Drought-associated chikungunya emergence along coastal East Africa. The American Journal of Tropical Medicine and Hygiene, 76(3), 405-407. https://doi.org/10.4269/ajtmh.2007.76.405 ↩
Cilloniz, C., Ewig, S., Gabarrus, A., Ferrer, M., Puig de la Bella Casa, J., Mensa, J., et Torres, A. (2017). Seasonality of pathogens causing community-acquired pneumonia. Respirology, 22(4), 778-785. https://doi.org/10.1111/resp.12978 ↩
Click Lambert, R., Kolivras, K. N., Resler, L. M., Brewster, C. C., et Paulson, S. L. (2008). The potential for emergence of Chagas disease in the United States. Geospatial Health, 2(2), 227-239. https://doi.org/10.4081/gh.2008.246 ↩
Clow, K. M., Leighton, P. A., Ogden, N. H., Lindsay, L. R., Michel, P., Pearl, D. L., et Jardine, C. M. (2017). Northward range expansion of Ixodes scapularis evident over a short timescale in Ontario, Canada. PLOS ONE, 12(12), 1-15. https://doi.org/10.1371/journal.pone.0189393 ↩
Collier, C. G. (1992). Weather conditions prior to major outbreaks of meningococcol meningitis in the United Kingdom. International Journal of Biometeorology, 36(1), 18-29. https://doi.org/10.1007/BF01208730 ↩
Comrie, A. C., et Glueck, M. F. (2007). Assessment of climate-coccidioidomycosis model: Model sensitivity for assessing climatologic effects on the risk of acquiring coccidioidomycosis. Annals of the New York Academy of Sciences, 1111(1), 83-95. https://doi.org/10.1196/annals.1406.024 ↩
Conza, L., Casati, S., Limoni, C., et Gaia, V. (2013). Meteorological factors and risk of community-acquired Legionnaires’ disease in Switzerland: An epidemiological study. BMJ Open, 3(3). https://doi.org/10.1136/bmjopen-2012-002428 ↩
Corrin, T., Greig, J., Harding, S., Young, I., Mascarenhas, M., et Waddell, L. A. (2018). Powassan virus, a scoping review of the global evidence. Zoonoses and Public Health, 65(6), 595-624. https://doi.org/10.1111/zph.12485 ↩
Cox, R., Sanchez, J., et Revie, C. W. (2013). Multi-criteria decision analysis tools for prioritising emerging or re-emerging infectious diseases associated with climate change in Canada. PLOS ONE, 8(8). https://doi.org/10.1371/journal.pone.0068338 ↩
Crampton, T. L., Light, R. B., Berg, G. M., Meyers, M. P., Schroeder, G. C., Hershfield, E. S., et Embil, J. M. (2002). Epidemiology and clinical spectrum of blastomycosis diagnosed at Manitoba hospitals. Clinical Infectious Diseases, 34(10), 1310-1316. https://doi.org/10.1086/340049 ↩
Cross, P. C., Edwards, W. H., Scurlock, B. M., Maichak, E. J., et Rogerson, J. D. (2007). Effects of management and climate on elk brucellosis in the Greater Yellowstone Ecosystem. Ecological Applications, 17(4), 957-964. https://doi.org/10.1890/06-1603 ↩
Dalcin, D., et Ahmed, S. Z. (2015). Blastomycosis in northwestern Ontario, 2004 to 2014. Canadian Journal of Infectious Diseases and Medical Microbiology, 26(5), 259-262. https://doi.org/10.1155/2015/468453 ↩
Davies, J. L., Epp, T., et Burgess, H. J. (2013). Prevalence and geographic distribution of canine and feline blastomycosis in the Canadian prairies. The Canadian Veterinary Journal, 54(8), 753-760. Consulté sur le site : https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3711163/ ↩
Davis, N. C. (1932). The effect of various temperatures in modifying the extrinsic incubation period of the yellow fever virus in Aëdes aegypti. American Journal of Epidemiology, 16(1), 163-176. https://doi.org/10.1093/oxfordjournals.aje.a117853 ↩
De Man, H., Mughini Gras, L., Schimmer, B., Friesema, I. H. M., De Roda Husman, A. M., et Van Pelt, W. (2016). Gastrointestinal, influenza-like illness and dermatological complaints following exposure to floodwater: A cross-sectional survey in the Netherlands. Epidemiology et Infection, 144(7), 1445-1454. https://doi.org/10.1017/S0950268815002654 ↩
DeFelice, N. B., Schneider, Z. D., Little, E., Barker, C., Caillouet, K. A., Campbell, S. R., . . . Shaman, J. (2018). Use of temperature to improve West Nile virus forecasts. PLoS computational biology, 14(3). https://doi.org/10.1371/journal.pcbi.1006047 ↩
Delisle, E., Rousseau, C., Broche, B., Leparc-Goffart, I., L’Ambert, G., Cochet, A., . . . Golliot, F. (2015). Chikungunya outbreak in Montpellier, France, September to October 2014. Eurosurveillance, 20(17), 1-6. https://doi.org/10.2807/1560-7917.ES2015.20.17.21108 ↩
Deplazes, P., Rinaldi, L., Alvarez Rojas, C. A., Torgerson, P. R., Harandi, M. F., Romig, T., . . . Jenkins, E. J. (2017). Global distribution of alveolar and cystic echinococcosis. Advances in Parasitology, 95, 315-493. https://doi.org/10.1016/bs.apar.2016.11.001 ↩
Dergousoff, S. J., Galloway, T. D., Lindsay, L. R., Curry, P. S., et Chilton, N. B. (2013). Range expansion of Dermacentor variabilis and Dermacentor andersoni (Acari: Ixodidae) near their northern distributional limits. Journal of Medical Entomology, 50(3), 510-520. https://doi.org/10.1603/me12193 ↩
DesGranges, J.-L., et Morneau, F. (2010). Potential sensitivity of Québec’s breeding birds to climate change. Avian Conservation and Ecology, 5(2). https://doi.org/10.5751/ace-00410-050205 ↩
Dibernardo, A., Cote, T., Ogden, N. H., et Lindsay, L. R. (2014). The prevalence of Borrelia miyamotoi infection, and co-infections with other Borrelia spp. in Ixodes scapularis ticks collected in Canada. Parasites et Vectors, 7, 1-8. https://doi.org/10.1186/1756-3305-7-183 ↩
Dicker, R. C., Coronado, F. t., Koo, D., et Gibson Parrish, R., II. (Eds.). (2012). Principles of epidemiology in public health practice: An introduction to applied epidemiology and biostatistics (3rd ed.). Atlanta, GA: Centers for Disease Control and Prevention.
↩
Dolcé, P., Bélanger, M.-J., Tumanowicz, K., Gauthier, C. P., Jutras, P., Massé, R., . . . Artsob, H. (2003). Coxiella burnetii seroprevalence of shepherds and their flocks in the lower Saint-Lawrence River region of Quebec, Canada. Canadian Journal of Infectious Diseases, 14(2), 97-102. https://doi.org/10.1155/2003/504796 ↩
Domenech de Cellès, M., Arduin, H., Lévy-Bruhl, D., Georges, S., Souty, C., Guillemot, D., . . . Opatowski, L. (2019). Unraveling the seasonal epidemiology of pneumococcus. Proceedings of the National Academy of Sciences of the United States of America, 116(5), 1802-1807. https://doi.org/10.1073/pnas.1812388116 ↩
Dowell, S. F. (2001). Seasonal variation in host susceptibility and cycles of certain infectious diseases. Emerging Infectious Diseases, 7(3), 369-374. https://doi.org/10.3201/eid0703.010301 ↩
Drebot, M. A., Artsob, H., et Werker, D. (2000). Hantavirus pulmonary syndrome in Canada, 1989-1999. Canada Communicable Disease Report, 26(8), 65-69. Consulté sur le site : http://publications.gc.ca/pub?id=9.507730etsl=0 ↩
Dworkin, M. S., Schwan, T. G., Anderson, D. E., et Borchardt, S. M. (2008). Tick-borne relapsing fever. Infectious Disease Clinics of North America, 22(3), 449-468. https://doi.org/10.1016/j.idc.2008.03.006 ↩
Dye, C. (2014). After 2015: Infectious diseases in a new era of health and development. Philosophical Transactions of the Royal Society B: Biological Sciences, 369(1645), 1-9. https://doi.org/10.1098/rstb.2013.0426 ↩
Eads, D. A., et Hoogland, J. L. (2017). Precipitation, climate change, and parasitism of prairie dogs by fleas that transmit plague. Journal of Parasitology, 103(4), 309-319. https://doi.org/10.1645/16-195 ↩
Earn, D. J. D., He, D., Loeb, M. B., Fonseca, K., Lee, B. E., et Dushoff, J. (2012). Effects of school closure on incidence of pandemic influenza in Alberta, Canada. Annals of Internal Medicine, 156(3), 173-181. https://doi.org/10.7326/0003-4819-156-3-201202070-00005 ↩
Earn, D. J. D., Ma, J., Poinar, H., Dushoff, J., Bolker, B. M. (2020). Acceleration of plague outbreaks in the second pandemic Proceedings of the National Academies of Science of the U S A, 117(44), 27703-27711.
↩
Ebi, K. L., Kovats, R. S., et Menne, B. (2006a). An approach for assessing human health vulnerability and public health interventions to adapt to climate change. Environmental Health Perspectives, 114(12), 1930-1934. https://doi.org/10.1289/ehp.8430 ↩
Ebi, K. L., Lewis, N. D., et Corvalan, C. (2006b). Climate variability and change and their potential health effects in small island states: Information for adaptation planning in the health sector. Environmental Health Perspectives, 114(12), 1957-1963. https://doi.org/10.1289/ehp.8429 ↩
Ebi, K. L., Ogden, N. H., Semenza, J. C., et Woodward, A. (2017). Detecting and attributing health burdens to climate change. Environmental Health Perspectives, 125(8), 1-8. https://doi.org/10.1289/EHP1509 ↩
Ebi, K.L., Balbus J.M., Luber G., Bole A., Crimmins A., Glass G., Saha S., Shimamoto M.M., Trtanj J., et White-Newsome J.L. (2018). Human Health. In Reidmiller, D.R., C.W. Avery, D.R. Easterling, K.E. Kunkel, K.L.M. Lewis, T.K. Maycock, and B.C. Stewart (Eds.), Impacts, Risks, and Adaptation in the United States: Fourth National Climate Assessment, Volume II (pp. 572–603). Washington, DC: U.S. Global Change Research Program. doi: 10.7930/NCA4.2018.CH14
↩
Edginton, S., Guan, T. H., Evans, G., et Srivastava, S. (2018). Human granulocytic anaplasmosis acquired from a blacklegged tick in Ontario. CMAJ, 190(12), E363-E366. https://doi.org/10.1503/CMAJ.171243 ↩
Eisen, R. J., Eisen, L., Ogden, N. H., et Beard, C. B. (2016). Linkages of weather and climate with Ixodes scapularis and Ixodes pacificus (Acari: Ixodidae), enzootic transmission of Borrelia burgdorferi, and Lyme disease in North America. Journal of Medical Entomology, 53(2), 250-261. https://doi.org/10.1093/jme/tjv199 ↩
Eldin, C., Mahamat, A., Djossou, F., et Raoult, D. (2015). Rainfall and sloth births in May, Q fever in July, Cayenne, French Guiana. The American Journal of Tropical Medicine and Hygiene, 92(5), 979-981. https://doi.org/10.4269/ajtmh.14-0751 ↩
Elrick, M. J., Gordon-Lipkin, E., Crawford, T. O., Van Haren, K., Messacar, K., Thornton, N., . . . Duggal, P. (2019). Clinical subpopulations in a sample of North American children diagnosed with acute flaccid myelitis, 2012-2016. JAMA Pediatrics, 173(2), 134-139. https://doi.org/10.1001/jamapediatrics.2018.4890 ↩
Embil, J. A., Tanner, C. E., Pereira, L. H., Staudt, M., Morrison, E. G., et Gualazzi, D. A. (1988). Seroepidemiologic survey of Toxocara canis infection in urban and rural children. Public Health, 102(2), 129-133. https://doi.org/10.1016/S0033-3506(88)80039-8 ↩
Enscore, R. E., Biggerstaff, B. J., Brown, T. L., Fulgham, R. E., Reynolds, P. J., Engelthaler, D. M., . . . Gage, K. L. (2002). Modeling relationships between climate and the frequency of human plague cases in the southwestern United States, 1960-1997. The American Journal of Tropical Medicine and Hygiene, 66(2), 186-196. https://doi.org/10.4269/ajtmh.2002.66.186 ↩
Escobar, L. E., Romero-Alvarez, D., Leon, R., Lepe-Lopez, M. A., Craft, M. E., Borbor-Cordova, M. J., et Svenning, J.-C. (2016). Declining prevalence of disease vectors under climate change. Scientific Reports, 6, 1-8. https://doi.org/10.1038/srep39150 ↩
Fields, B. S., Benson, R. F., et Besser, R. E. (2002). Legionella and Legionnaires’ disease: 25 years of investigation. Clinical Microbiology Reviews, 15(3), 506-526. https://doi.org/10.1128/cmr.15.3.506-526.2002 ↩
Findlater, A., et Bogoch, I. I. (2018). Human mobility and the global spread of infectious diseases: A focus on air travel. Trends in Parasitology, 34(9), 772-783. https://doi.org/10.1016/j.pt.2018.07.004 ↩
Fisman, D. N., Lim, S., Wellenius, G. A., Johnson, C., Britz, P., Gaskins, M., . . . Newbern, C. (2005). It’s not the heat, it’s the humidity: Wet weather increases legionellosis risk in the greater Philadelphia metropolitan area. The Journal of Infectious Diseases, 192(12), 2066-2073. https://doi.org/10.1086/498248 ↩
Fisman, D. N., Tuite, A. R., et Brown, K. A. (2016). Impact of El Niño Southern Oscillation on infectious disease hospitalization risk in the United States. Proceedings of the National Academy of Sciences of the United States of America, 113(51), 14589-14594. https://doi.org/10.1073/pnas.1604980113 ↩
Fitzhenry, R., Weiss, D., Cimini, D., Balter, S., Boyd, C., Alleyne, L., . . . Varma, J. K. (2017). Legionnaires’ disease outbreaks and cooling towers, New York City, New York, USA. Emerging Infectious Diseases, 23(11), 1769-1776. https://doi.org/10.3201/eid2311.161584 ↩
Fonseca, K., Prince, G. D., Bratvold, J., Fox, J. D., Pybus, M., Preksaitis, J. K., et Tilley, P. (2005). West Nile virus infection and conjunctival exposure. Emerging Infectious Diseases, 11(10), 1648-1649. https://doi.org/10.3201/eid1110.040212 ↩
Forbes, L. B. (1991). Isolates of Brucella suis biovar 4 from animals and humans in Canada, 1982-1990. The Canadian Veterinary Journal, 32(11), 686-688. Consulté sur le site : https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1481085/ ↩
Franco-Paredes, C., Chastain, D., Taylor, P., Stocking, S., et Sellers, B. (2017). Boar hunting and brucellosis caused by Brucella suis. Travel Med Infect Dis, 16, 18-22. https://doi.org/10.1016/j.tmaid.2017.03.006 ↩
Friel, S. (2019). Climate change and the people’s health (N. Krieger, Ed.). Oxford University Press. doi:10.1093/oso/9780190492731.001.0001
↩
Gabriele-Rivet, V., Arsenault, J., Badcock, J., Cheng, A., Edsall, J., Goltz, J., . . . Ogden, N. H. (2015). Different ecological niches for ticks of public health significance in Canada. PLOS ONE, 10(7), 1-19. https://doi.org/10.1371/journal.pone.0131282 ↩
Gallant, D., Slough, B. G., Reid, D. G., et Berteaux, D. (2012). Arctic fox versus red fox in the warming Arctic: Four decades of den surveys in north Yukon. Polar Biology, 35(9), 1421-1431. https://doi.org/10.1007/s00300-012-1181-8 ↩
Garcia-Solache, M. A., et Casadevall, A. (2010). Global warming will bring new fungal diseases for mammals. mBio, 1(1), 1-3. https://doi.org/10.1128/mBio.00061-10 ↩
Garcia-Vidal, C., Labori, M., Viasus, D., Simonetti, A., Garcia-Somoza, D., Dorca, J., . . . Carratalà, J. (2013). Rainfall is a risk factor for sporadic cases of Legionella pneumophila pneumonia. PLOS ONE, 8(4), 1-5. https://doi.org/10.1371/journal.pone.0061036 ↩
Gargan, T. P., II., Clark, G. G., Dohm, D. J., Turell, M. J., et Bailey, C. L. (1988). Vector potential of selected North American mosquito species for Rift Valley fever virus. The American Journal of Tropical Medicine and Hygiene, 38(2), 440-446. https://doi.org/10.4269/ajtmh.1988.38.440 ↩
Garroway, C. J., Bowman, J., Cascaden, T. J., Holloway, G. L., Mahan, C. G., Malcolm, J. R., . . . Wilson, P. J. (2010). Climate change induced hybridization in flying squirrels. Global Change Biology, 16(1), 113-121. https://doi.org/10.1111/j.1365-2486.2009.01948.x ↩
Garroway, C. J., Bowman, J., Holloway, G. L., Malcolm, J. R., et Wilson, P. J. (2011). The genetic signature of rapid range expansion by flying squirrels in response to contemporary climate warming. Global Change Biology, 17(5), 1760-1769. https://doi.org/10.1111/j.1365-2486.2010.02384.x ↩
Gasmi, S., Bouchard, C., Ogden, N. H., Adam-Poupart, A., Pelcat, Y., Rees, E. E., . . . Thivierge, K. (2018). Evidence for increasing densities and geographic ranges of tick species of public health significance other than Ixodes scapularis in Québec, Canada. PLOS ONE, 13(8), 1-15. https://doi.org/10.1371/JOURNAL.PONE.0201924 ↩
Gasmi, S., Ogden, N. H., Lindsay, L. R., Burns, S., Fleming, S., Badcock, J., . . . Koffi, J. K. (2017). Surveillance for Lyme disease in Canada: 2009-2015. Canada Communicable Disease Report, 43(10), 194-199. https://doi.org/10.14745/ccdr.v43i10a01 ↩
Gilbert, M., Slingenbergh, J., et Xiao, X. (2008). Climate change and avian influenza. OIE Scientific and Technical Review, 27(2), 459-466. https://doi.org/10.20506/rst.27.2.1821 ↩
Gilmour, M. W., Graham, M., Reimer, A., et Van Domselaar, G. (2013). Public health genomics and the new molecular epidemiology of bacterial pathogens. Public Health Genomics, 16(1-2), 25-30. https://doi.org/10.1159/000342709 ↩
Giordano, B. V., Gasparotto, A., et Hunter, F. F. (2015). A checklist of the 67 mosquito species of Ontario, Canada. Journal of the American Mosquito Control Association, 31(1), 101-103. https://doi.org/10.2987/14-6456r.1 ↩
Giordano, B. V., Kaur, S., et Hunter, F. F. (2017). West Nile virus in Ontario, Canada: A twelve-year analysis of human case prevalence, mosquito surveillance, and climate data. PLOS ONE, 12(8), 1-15. https://doi.org/10.1371/journal.pone.0183568 ↩
Giordano, B. V., Turner, K. W., et Hunter, F. F. (2018). Geospatial analysis and seasonal distribution of West Nile virus vectors (Diptera: Culicidae) in Southern Ontario, Canada. International Journal of Environmental Research and Public Health, 15(4), 1-18. https://doi.org/10.3390/ijerph15040614 ↩
Githeko, A. K., Lindsay, S. W., Confalonieri, U. E., et Patz, J. A. (2000). Climate change and vector-borne diseases: A regional analysis. Bulletin of the World Health Organization, 78(9), 1136-1147. Consulté sur le site : https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2560843/ ↩
Gleason, J. A., Kratz, N. R., Greeley, R. D., et Fagliano, J. A. (2016). Under the weather: Legionellosis and meteorological factors. EcoHealth, 13(2), 293-302. https://doi.org/10.1007/s10393-016-1115-y ↩
Global Burden of Disease 2015 Mortality and Causes of Death Collaborators. (2016). Global, regional, and national life expectancy, all-cause mortality, and cause-specific mortality for 249 causes of death, 1980–2015: A systematic analysis for the Global Burden of Disease Study 2015. The Lancet, 388(10053), 1459-1544. https://doi.org/10.1016/S0140-6736(16)31012-1 ↩
Golnar, A. J., Kading, R. C., et Hamer, G. L. (2018). Quantifying the potential pathways and locations of Rift Valley fever virus entry into the United States. Transboundary and Emerging Diseases, 65(1), 85-95. https://doi.org/10.1111/tbed.12608 ↩
Golnar, A. J., Turell, M. J., LaBeaud, A. D., Kading, R. C., et Hamer, G. L. (2014). Predicting the mosquito species and vertebrate species involved in the theoretical transmission of Rift Valley fever virus in the United States. PLOS Neglected Tropical Diseases, 8(9), 1-11. https://doi.org/10.1371/journal.pntd.0003163 ↩
Goneau, L. W., Mehta, K., Wong, J., L’Huillier, A. G., et Gubbay, J. B. (2018). Zoonotic influenza and human health — Part 1: Virology and epidemiology of zoonotic influenzas. Current Infectious Disease Reports, 20(10), 1-12. https://doi.org/10.1007/s11908-018-0642-9 ↩
Gorris, M. E., Cat, L. A., Zender, C. S., Treseder, K. K., et Randerson, J. T. (2018). Coccidioidomycosis dynamics in relation to climate in the southwestern United States. GeoHealth, 2(1), 6-24. https://doi.org/10.1002/2017GH000095 ↩
Gouvernement du Manitoba. (2019b). Manitoba Health, Seniors and Active Living West Nile Virus Program 2019: Planning Documents for Municipalities III. Adult Mosquito Control Package. Consulté sur le site : https://www.gov.mb.ca/health/wnv/docs/amcontrolontrolpackage.pdf ↩
Granados, D. P., et Castañeda, E. (2006). Influence of climatic conditions on the isolation of members of the Cryptococcus neoformans species complex from trees in Colombia from 1992-2004. FEMS Yeast Research, 6(4), 636-644. https://doi.org/10.1111/j.1567-1364.2006.00090.x ↩
Greer, A. L., Drews, S. J., et Fisman, D. N. (2009). Why « winter » vomiting disease? Seasonality, hydrology, and norovirus epidemiology in Toronto, Canada. EcoHealth, 6(2), 192-199. https://doi.org/10.1007/s10393-009-0247-8 ↩
Greer, A., Ng, V., et Fisman, D. (2008). Climate change and infectious diseases in North America: The road ahead. CMAJ, 178(6), 715-722. https://doi.org/10.1503/cmaj.081325 ↩
Griot, C., Gonzalez-Scarano, F., et Nathanson, N. (1993). Molecular determinants of the virulence and infectivity of California serogroup bunyaviruses. Annual Review of Microbiology, 47, 117-138. https://doi.org/10.1146/annurev.mi.47.100193.001001 ↩
Groupe d’experts intergouvernemental sur l’évolution du climat (GIEC). (2014). Résumé à l’intention des décideurs. Sous la direction de Field, C.B., V.R. Barros, D.J. Dokken, K.J. Mach, M.D. Mastrandrea, T.E. Bilir, M. Chatterjee, K.L. Ebi, Y.O. Estrada, R.C. Genova, B. Girma, E.S. Kissel, A.N. Levy, S. MacCracken, P.R. Mastrandrea, and L.L. White (éds.), Changements climatiques 2014: Incidences, adaptation et vulnérabilité. Part A: Global and Sectoral Aspects.Contribution du Groupe de travail II au cinquième Rapport d’évaluation du Groupe d’experts intergouvernemental sur l’évolution du climat. Cambridge, Royaume-Uni and New York, NY, ÉUA: Cambridge University Press.
↩
Groupe de recherche en épidémiologie des zoonoses et santé publique. (2019). Consulté sur le site : https://grezosp.com/ ↩
Gubler, D. J. (2007). The continuing spread of West Nile virus in the Western Hemisphere. Clinical Infectious Diseases, 45(8), 1039-1046. https://doi.org/10.1086/521911 ↩
Guerra, M., Walker, E. D., Jones, C., Paskewitz, S., Cortinas, M. R., Stancil, A., . . . Kitron, U. (2002). Predicting the risk of Lyme disease: Habitat suitability for Ixodes scapularis in the North Central United States. Emerging Infectious Diseases, 8(3), 289-297. Consulté sur le site : https://wwwnc.cdc.gov/eid/article/8/3/01-0166_article ↩
Gupta, S., Parker, J., Smits, S., Underwood, J., et Dolwani, S. (2020). Persistent viral shedding of SARS-CoV-2 in faeces – a rapid review. Colorectal disease: the official journal of the Association of Coloproctology of Great Britain and Ireland, 22(6), 611–620. https://doi.org/10.1111/codi.15138 ↩
Hajizadeh M, Mitnitski A, Rockwood K. 2016. Socioeconomic gradient in health in Canada: Is the gap widening or narrowing? Health Policy, 120(9), 1040-50. https://doi.org/10.1016/j.healthpol.2016.07.019.
↩
Halsby, K. D., Joseph, C. A., Lee, J. V., et Wilkinson, P. (2014). The relationship between meteorological variables and sporadic cases of Legionnaires’ disease in residents of England and Wales. Epidemiology et Infection, 142(11), 2352-2359. https://doi.org/10.1017/S0950268813003294 ↩
Harrigan, R. J., Thomassen, H. A., Buermann, W., et Smith, T. B. (2014). A continental risk assessment of West Nile virus under climate change. Global Change Biology, 20(8), 2417-2425. https://doi.org/10.1111/gcb.12534 ↩
Hassel, C., Mirand, A., Lukashev, A., Terletskaia-Ladwig, E., Farkas, A., Schuffenecker, I., . . . Bailly, J.-L. (2015). Transmission patterns of human enterovirus 71 to, from and among European countries, 2003 to 2013. Eurosurveillance, 20(34), 1-11. https://doi.org/10.2807/1560-7917.ES.2015.20.34.30005 ↩
Heesterbeek, H., Anderson, R. M., Andreasen, V., Bansal, S., De Angelis, D., Dye, C., . . . Isaac Newton Institute IDD Collaboration. (2015). Modeling infectious disease dynamics in the complex landscape of global health. Science, 347(6227), 1-10. https://doi.org/10.1126/science.aaa4339 ↩
Henao-Restrepo, A. M., Camacho, A., Longini, I. M., Watson, C. H., Edmunds, W. J., Egger, M., . . . Kieny, M.-P. (2017). Efficacy and effectiveness of an rVSV-vectored vaccine in preventing Ebola virus disease: Final results from the Guinea ring vaccination, open-label, cluster-randomised trial (Ebola Ça Suffit!). The Lancet, 389(10068), 505-518. https://doi.org/10.1016/S0140-6736(16)32621-6 ↩
Henri, D.A., Jean-Gagnon, F., et Grant Gilchrist, H. (2018). Using Inuit traditional ecological knowledge for detecting and monitoring avian cholera among Common Eiders in the eastern Canadian Arctic. Ecology and Society, 23(1), 22.
↩
Hersteinsson, P., et MacDonald, D. W. (1992). Interspecific competition and the geographical distribution of red and arctic foxes Vulpes vulpes and Alopex lagopus. Oikos, 64(3), 505-515. https://doi.org/10.2307/3545168 ↩
Hertig, E. (2019). Distribution of Anopheles vectors and potential malaria transmission stability in Europe and the Mediterranean area under future climate change. Parasites et Vectors, 12, 1-9. https://doi.org/10.1186/s13071-018-3278-6 ↩
Hicks, L. A., Rose, C. E., Fields, B. S., Drees, M. L., Engel, J. P., Jenkins, P. R., . . . Whitney, C. G. (2007). Increased rainfall is associated with increased risk for legionellosis. Epidemiology et Infection, 135(5), 811-817. https://doi.org/10.1017/S0950268806007552 ↩
Hierlihy, C., Waddell, L., Young, I., Greig, J., Corrin, T., et Mascarenhas, M. (2019). A systematic review of individual and community mitigation measures for prevention and control of chikungunya virus. PLOS ONE, 14(2), 1-27. https://doi.org/10.1371/journal.pone.0212054 ↩
Hill A.B. (1965). The environment and disease: association or causation? Proceedings of the Royal Society of Medicine, 58(5), 295–300.
↩
Hitch, A. T., et Leberg, P. L. (2007). Breeding distributions of North American bird species moving north as a result of climate change. Conservation Biology, 21(2), 534-539. https://doi.org/10.1111/j.1523-1739.2006.00609.x ↩
Hoegh-Guldberg, O., Jacob, D., Taylor, M., Bindi, M., Brown, S., Camilloni, I., . . . Zhou, G. (2019). Impacts of 1.5°C of global warming on natural and human systems. In V. Masson-Delmotte, P. Zhai, H.-O. Pörtner, D. Roberts, J. Skea, P. R. Shukla, A. Pirani, W. Moufouma-Okia, C. Péan, R. Pidcock, S. Connors, J. B. R. Matthews, Y. Chen, X. Zhou, M. I. Gomis, E. Lonnoy, T. Maycock, M. Tignor, et T. Waterfield (Eds.), Global warming of 1.5°C: An IPCC special report. In Press. Consulté sur le site : https://www.ipcc.ch/sr15/chapter/chapter-3/ ↩
Holm-Hansen, C. C., Midgley, S. E., et Fischer, T. K. (2016). Global emergence of enterovirus D68: a systematic review. The Lancet Infectious Diseases, 16(5), e64-e75. https://doi.org/10.1016/S1473-3099(15)00543-5 ↩
Holt, A. C., Salkeld, D. J., Fritz, C. L., Tucker, J. R., et Gong, P. (2009). Spatial analysis of plague in California: Niche modeling predictions of the current distribution and potential response to climate change. International Journal of Health Geographics, 8, 1-14. https://doi.org/10.1186/1476-072x-8-38 ↩
Hongoh, V., Berrang-Ford, L., Scott, M. E., et Lindsay, L. R. (2012). Expanding geographical distribution of the mosquito, Culex pipiens, in Canada under climate change. Applied Geography, 33, 53-62. https://doi.org/10.1016/j.apgeog.2011.05.015 ↩
Huettmann, F., Magnuson, E. E., et Hueffer, K. (2017). Ecological niche modeling of rabies in the changing Arctic of Alaska. Acta Veterinaria Scandinavica, 59, 1-11. https://doi.org/10.1186/s13028-017-0285-0 ↩
Hunter, M., Donnelly, C., Smart, D., Smyth, B., Menzies, F., et Hedderwick, S. (2015). Brucellosis in people with occupational cattle exposure in Northern Ireland: Clinical features of 53 cases. Journal of Infection, 70(1), 101-103. https://doi.org/10.1016/j.jinf.2014.08.004 ↩
Institut national de santé publique du Québec. (2018a). Observatoire multipartite québécois sur les zoonoses et l’adaptation aux changements climatiques. Consulté sur le site : https://www.inspq.qc.ca/zoonoses/observatoire ↩
Institut national de santé publique du Québec. (2018b). Priorisation des zoonoses au Québec dans un contexte d’adaptation aux changements climatiques à l’aide d’un outil d’aide à la décision multicritère. Consulté sur le site : https://www.inspq.qc.ca/publications/2432 ↩
Iranpour, M., Hebert, P., Dibernardo, A., et Lindsay, R. (2017). Ochlerotatus ventrovittis Dyar (Diptera: Culicidae): A new record for the mosquito fauna of Canada. Paper presented at the Joint Meeting of the Entomological Societies of Canada and Manitoba: October 22-25, 2017, Winnipeg, MB. Consulté sur le site : https://home.cc.umanitoba.ca/~fieldspg/pdf/Proceedings/ESMproceedings_V73.pdf ↩
Iranpour, M., Turell, M. J., et Lindsay, L. R. (2011). Potential for Canadian mosquitoes to transmit Rift Valley fever virus. Journal of the American Mosquito Control Association, 27(4), 363-369. https://doi.org/10.2987/11-6169.1 ↩
Jenkins, E. J., Castrodale, L. J., de Rosemond, S. J. C., Dixon, B. R., Elmore, S. A., Gesy, K. M., . . . Thompson, R. C. A. (2013). Tradition and transition: Parasitic zoonoses of people and animals in Alaska, northern Canada, and Greenland. Advances in Parasitology, 82, 33-204. https://doi.org/10.1016/B978-0-12-407706-5.00002-2 ↩
Jenkins, E. J., Schurer, J. M., et Gesy, K. M. (2011). Old problems on a new playing field: Helminth zoonoses transmitted among dogs, wildlife, and people in a changing northern climate. Veterinary Parasitology, 182(1), 54-69. https://doi.org/10.1016/j.vetpar.2011.07.015 ↩
Jetten, T. H., et Focks, D. A. (1997). Potential changes in the distribution of dengue transmission under climate warming. The American Journal of Tropical Medicine and Hygiene, 57(3), 285-297. https://doi.org/10.4269/ajtmh.1997.57.285 ↩
Ji, W., Huh, K., Kang, M., Hong, J., Bae, G. H., Lee, R., Na, Y., Choi, H., Gong, S. Y., Choi, Y. H., Ko, K. P., Im, J. S., et Jung, J. (2020). Effect of Underlying Comorbidities on the Infection and Severity of COVID-19 in Korea: a Nationwide Case-Control Study. Journal of Korean medical science, 35(25), e237. https://doi.org/10.3346/jkms.2020.35.e237 ↩
Jones, B., et O’Neill, B. C. (2016). Spatially explicit global population scenarios consistent with the Shared Socioeconomic Pathways. Environmental Research Letters, 11(8), 1-10. https://doi.org/10.1088/1748-9326/11/8/084003 ↩
Jones, K. E., Patel, N. G., Levy, M. A., Storeygard, A., Balk, D., Gittleman, J. L., et Daszak, P. (2008). Global trends in emerging infectious diseases. Nature, 451(7181), 990-993. https://doi.org/10.1038/nature06536 ↩
Jüni, P., Rothenbühler, M., Bobos, P., Thorpe, K. E., da Costa, B. R., Fisman, D. N., Slutsky, A. S., et Gesink, D. (2020). Impact of climate and public health interventions on the COVID-19 pandemic: a prospective cohort study. CMAJ : Canadian Medical Association journal = journal de l’Association medicale canadienne, 192(21), E566–E573. https://doi.org/10.1503/cmaj.200920 ↩
Kamal, M., Kenawy, M. A., Rady, M. H., Khaled, A. S., et Samy, A. M. (2019). Mapping the global potential distributions of two arboviral vectors Aedes aegypti and Ae. albopictus under changing climate. PLOS ONE, 13(12), 1-21. https://doi.org/10.1371/journal.pone.0210122 ↩
Kanzawa, M., Spindler, H., Anglemyer, A., et Rutherford, G. W. (2020). Will Coronavirus Disease 2019 Become Seasonal? The Journal of infectious diseases, 222(5), 719–721. https://doi.org/10.1093/infdis/jiaa345 ↩
Kauffman, C. A. (2007). Histoplasmosis: A clinical and laboratory update. Clinical Microbiology Reviews, 20(1), 115-132. https://doi.org/10.1128/CMR.00027-06 ↩
Kausrud, K. L., Begon, M., Ari, T. B., Viljugrein. H., Esper, J., Büntgen, U., Leirs, H., Junge, C., Yang, B., Yang, M., Xu, L., Stenseth, N. C. (2010) Modeling the epidemiological history of plague in Central Asia: palaeoclimatic forcing on a disease system over the past millennium. BMC Biology, 8, 112. https://doi.org/10.1186/1741-7007-8-112.
↩
Kausrud, K. L., Mysterud, A., Steen, H., Vik, J. O., Østbye, E., Cazelles, B., . . . Stenseth, N. C. (2008). Linking climate change to lemming cycles. Nature, 456(7218), 93-97. https://doi.org/10.1038/nature07442 ↩
KC, S., et Lutz, W. (2017). The human core of the shared socioeconomic pathways: Population scenarios by age, sex and level of education for all countries to 2100. Global Environmental Change, 42, 181-192. https://doi.org/10.1016/j.gloenvcha.2014.06.004 ↩
Keegan, J. D., et Holland, C. V. (2010). Contamination of the hair of owned dogs with the eggs of Toxocara spp. Veterinary Parasitology, 173(1-2), 161-164. https://doi.org/10.1016/j.vetpar.2010.06.010 ↩
Kepron, M. W., Schoemperlen, C. B., Hershfield, E. S., Zylak, C. J., et Cherniack, R. M. (1972). North American blastomycosis in Central Canada: A review of 36 cases. CMAJ, 106(3), 243-246. Consulté sur le site : https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1940364/ ↩
Kidd, S. E., Bach, P. J., Hingston, A. O., Mak, S., Chow, Y., MacDougall, L., . . . Bartlett, K. H. (2007a). Cryptococcus gattii dispersal mechanisms, British Columbia, Canada. Emerging Infectious Diseases, 13(1), 51-57. https://doi.org/10.3201/eid1301.060823 ↩
Kidd, S. E., Chow, Y., Mak, S., Bach, P. J., Chen, H., Hingston, A. O., . . . Bartlett, K. H. (2007b). Characterization of environmental sources of the human and animal pathogen Cryptococcus gattii in British Columbia, Canada, and the Pacific Northwest of the United States. Applied and Environmental Microbiology, 73(5), 1433-1443. https://doi.org/10.1128/AEM.01330-06 ↩
Kidd, S. E., Hagen, F., Tscharke, R. L., Huynh, M., Bartlett, K. H., Fyfe, M., Macdougall, L., Boekhout, T., Kwon-Chung, K. J., et Meyer, W. (2004). A rare genotype of Cryptococcus gattii caused the cryptococcosis outbreak on Vancouver Island (British Columbia, Canada). Proceedings of the National Academy of Sciences of the United States of America, 101(49), 17258–17263. https://doi.org/10.1073/pnas.0402981101 ↩
Kilpatrick, A. M., Dobson, A. D. M., Levi, T., Salkeld, D. J., Swei, A., Ginsberg, H. S., . . . Diuk-Wasser, M. A. (2017). Lyme disease ecology in a changing world: Consensus, uncertainty and critical gaps for improving control. Philosophical Transactions of the Royal Society B: Biological Sciences, 372(1722). https://doi.org/10.1098/rstb.2016.0117 ↩
Kilpatrick, A. M., Kramer, L. D., Jones, M. J., Marra, P. P., et Daszak, P. (2006). West Nile virus epidemics in North America are driven by shifts in mosquito feeding behavior. PLOS Biology, 4(4), 0606-0610. https://doi.org/10.1371/journal.pbio.0040082 ↩
Kim, B. I., Ki, H., Park, S., Cho, E., et Chun, B. C. (2016). Effect of climatic factors on hand, foot, and mouth disease in South Korea, 2010-2013. PLOS ONE, 11(6), 1-9. https://doi.org/10.1371/journal.pone.0157500 ↩
Kinlin, L. M., Spain, C. V., Ng, V., Johnson, C. C., White, A. N. J., et Fisman, D. N. (2009). Environmental exposures and invasive meningococcal disease: An evaluation of effects on varying time scales. American Journal of Epidemiology, 169(5), 588-595. https://doi.org/10.1093/aje/kwn383 ↩
Kissler, S. M., Tedijanto, C., Goldstein, E., Grad, Y. H., et Lipsitch, M. (2020). Projecting the transmission dynamics of SARS-CoV-2 through the postpandemic period. Science (New York, N.Y.), 368(6493), 860–868. https://doi.org/10.1126/science.abb5793 ↩
Knobel, D. L., Cleaveland, S., Coleman, P. G., Fèvre, E. M., Meltzer, M. I., Miranda, M. E. G., . . . Meslin, F.-X. (2005). Re-evaluating the burden of rabies in Africa and Asia. Bulletin of the World Health Organization, 83(5), 360-368. Consulté sur le site : https://www.who.int/bulletin/volumes/83/5/360.pdf ↩
Kotchi S.-O., Bouchard, C., Brazeau, S., Ogden, N.H. (2021) Earth Observation-informed risk maps of the Lyme disease vector Ixodes scapularis in Central and Eastern Canada. Remote Sensing, 13(3), 524.
↩
Kotwa, J. D., Isaksson, M., Jardine, C. M., Campbell, G. D., Berke, O., Pearl, D. L., . . . Peregrine, A. S. (2019). Echinococcus multilocularis infection, Southern Ontario, Canada. Emerging Infectious Diseases, 25(2), 265-272. https://doi.org/10.3201/eid2502.180299 ↩
Kraemer, M. U. G., Reiner, R. C., Jr., Brady, O. J., Messina, J. P., Gilbert, M., Pigott, D. M., . . . Golding, N. (2019). Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopictus. Nature Microbiology, 4, 854-863. https://doi.org/10.1038/s41564-019-0376-y ↩
Kraemer, M. U. G., Sinka, M. E., Duda, K. A., Mylne, A., Shearer, F. M., Brady, O. J., . . . Hay, S. I. (2015). The global compendium of Aedes aegypti and Ae. albopictus occurrence. Scientific Data, 2, 1-8. https://doi.org/10.1038/sdata.2015.35 ↩
Kulkarni, M. A., Berrang-Ford, L., Buck, P. A., Drebot, M. A., Lindsay, L. R., et Ogden, N. H. (2015). Major emerging vector-borne zoonotic diseases of public health importance in Canada. Emerging Microbes et Infections, 4(1), 1-7. https://doi.org/10.1038/emi.2015.33 ↩
Kuster, S. P., Tuite, A. R., Kwong, J. C., McGeer, A., The Toronto Invasive Bacterial Diseases Network, et Fisman, D. N. (2011). Evaluation of coseasonality of influenza and invasive pneumococcal disease: Results from prospective surveillance. PLOS Medicine, 8(6), 1-9. https://doi.org/10.1371/journal.pmed.1001042 ↩
Kwon-Chung, K. J., et Bennett, J. E. (1984). High prevalence of Cryptococcus neoformans var. gattii in tropical and subtropical regions. Zentralblatt für Bakteriologie, Mikrobiologie, und Hygiene Serie A – Medical microbiology, infectious diseases, virology, parasitology, 257(2), 213-218.
↩
Larrieu, S., Cassadou, S., Rosine, J., Chappert, J. L., Blateau, A., Ledrans, M., et Quénel, P. (2014). Lessons raised by the major 2010 dengue epidemics in the French West Indies. Acta Tropica, 131, 37-40. https://doi.org/10.1016/j.actatropica.2013.11.023 ↩
Larsen, J. N., Anisimov, O. A., Constable, A., Hollowed, A. B., Maynard, N., Prestrud, P., . . . Stone, J. M. R. (2014). Polar regions. In V. R. Barros, C. B. Field, D. J. Dokken, M. D. Mastrandrea, K. J. Mach, T. E. Bilir, M. Chatterjee, K. L. Ebi, Y. O. Estrada, R. C. Genova, B. Girma, E. S. Kissel, A. N. Levy, S. MacCracken, P. R. Mastrandrea, et L. L. White (Eds.), Climate change 2014: Impacts, adaptation, and vulnerability. Part B: Regional aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (pp. 1567-1612). Cambridge, United Kingdom: Cambridge University Press. Consulté sur le site : https://www.ipcc.ch/report/ar5/wg2/ ↩
Lau, L., Wudel, B., Kadkhoda, K., et Keynan, Y. (2017). Snowshoe hare virus causing meningoencephalitis in a young adult from northern Manitoba, Canada. Open Forum Infectious Diseases, 4(3), 1-3. https://doi.org/10.1093/ofid/ofx150 ↩
Lau, S., Luk, H., Wong, A., Li, K., Zhu, L., He, Z., Fung, J., Chan, T., Fung, K., et Woo, P. (2020). Possible Bat Origin of Severe Acute Respiratory Syndrome Coronavirus 2. Emerging infectious diseases, 26(7), 1542–1547. https://doi.org/10.3201/eid2607.200092 ↩
Lee, A. C. Y., Schantz, P. M., Kazacos, K. R., Montgomery, S. P., et Bowman, D. D. (2010). Epidemiologic and zoonotic aspects of ascarid infections in dogs and cats. Trends in Parasitology, 26(4), 155-161. https://doi.org/10.1016/j.pt.2010.01.002 ↩
Leighton, P. A., Koffi, J. K., Pelcat, Y., Lindsay, L. R., et Ogden, N. H. (2012). Predicting the speed of tick invasion: An empirical model of range expansion for the Lyme disease vector Ixodes scapularis in Canada. Journal of Applied Ecology, 49(2), 457-464. https://doi.org/10.1111/j.1365-2664.2012.02112.x ↩
Levac, D., Colquhoun, H., et O’Brien, K. K. (2010). Scoping studies: Advancing the methodology. Implementation Science, 5, 69. https://doi.org/10.1186/1748-5908-5-69 ↩
Levine RS, Hedeen DL, Hedeen MW, Hamer GL, Mead DG, Kitron UD. (2017). Avian species diversity and transmission of West Nile virus in Atlanta, Georgia. Parasites et Vectors, 10(1), 62. https://doi.org/10.1186/s13071-017-1999-6.
↩
Liao, J., Yu, S., Yang, F., Yang, M., Hu, Y., et Zhang, J. (2016). Short-term effects of climatic variables on hand, foot, and mouth disease in mainland China, 2008-2013: A multilevel spatial Poisson regression model accounting for overdispersion. PLOS ONE, 11(1), 1-14. https://doi.org/10.1371/journal.pone.0147054 ↩
Light, R. B., Kralt, D., Embil, J. M., Trepman, E., Wiebe, L., Limerick, B., . . . Macdonald, K. (2008). Seasonal variations in the clinical presentation of pulmonary and extrapulmonary blastomycosis. Medical Mycology, 46(8), 835-841. https://doi.org/10.1080/13693780802132763 ↩
Likos, A., Griffin, I., Bingham, A. M., Stanek, D., Fischer, M., White, S., . . . Philip, C. (2016). Local mosquito-borne transmission of Zika virus—Miami-Dade and Broward counties, Florida, June–August 2016. Morbidity and Mortality Weekly Report, 65(38), 1032-1038. https://doi.org/10.15585/mmwr.mm6538e1 ↩
Lin, H., Zou, H., Wang, Q., Liu, C., Lang, L., Hou, X., et Li, Z. (2013). Short-term effect of El Niño-Southern Oscillation on pediatric hand, foot and mouth disease in Shenzhen, China. PLOS ONE, 8(7), 1-7. http://doi.org/10.1371/journal.pone.0065585 ↩
Lindquist, E. E., Galloway, T. D., Artsob, H., Lindsay, L. R., Drebot, M., Wood, H., et Robbins, R. G. (2016). A handbook to the ticks of Canada (Ixodida: Ixodidae, Argasidae): Biological Survey of Canada Monograph Series No. 7: Biological Survey of Canada. Consulté sur le site : https://biologicalsurvey.ca/public/Bsc/Controller/Page/AGR-001-Ticks-Monograph.pdf ↩
Lindsay, A. P., Hope, V., Marshall, R. J., et Salinger, J. (2002). Meningococcal disease and meteorological conditions in Auckland, New Zealand. Australian and New Zealand Journal of Public Health, 26(3), 212-218. https://doi.org/10.1111/j.1467-842X.2002.tb00676.x ↩
Lindsay, L. R., Barker, I. K., Surgeoner, G. A., Mcewen, S. A., Gillespie, T. J., et Addison, E. M. (1998). Survival and development of the different life stages of Ixodes scapularis (Acari: Ixodidae) held within four habitats on Long Point, Ontario, Canada. Journal of Medical Entomology, 35(3), 189-199. https://doi.org/10.1093/jmedent/35.3.189 ↩
Lindsey, N. P., Staples, J. E., et Fischer, M. (2018). Chikungunya virus disease among travelers—United States, 2014-2016. The American Journal of Tropical Medicine and Hygiene, 98(1), 192-197. https://doi.org/10.4269/ajtmh.17-0668 ↩
Litvinjenko, S., et Lunny, D. (2017). Blastomycosis hospitalizations in northwestern Ontario: 2006-2015. Canada Communicable Disease Report, 43(10), 200-205. https://doi.org/10.14745/ccdr.v43i10a02 ↩
Litvinov, I. V., St-Germain, G., Pelletier, R., Paradis, M., et Sheppard, D. C. (2013). Endemic human blastomycosis in Quebec, Canada, 1988–2011. Epidemiology et Infection, 141(6), 1143-1147. https://doi.org/10.1017/S0950268812001860 ↩
Litvintseva, A. P., Marsden-Haug, N., Hurst, S., Hill, H., Gade, L., Driebe, E. M., . . . Chiller, T. (2015). Valley fever: Finding new places for an old disease: Coccidioides immitis found in Washington State soil associated with recent human infection. Clinical Infectious Diseases, 60(1), e1-e3. https://doi.org/10.1093/cid/ciu681 ↩
Lohrenz, S., Minion, J., Pandey, M., et Karunakaran, K. (2018). Blastomycosis in southern Saskatchewan 2000–2015: Unique presentations and disease characteristics. Medical Mycology, 56(7), 787-795. https://doi.org/10.1093/mmy/myx131 ↩
Lopman, B., Armstrong, B., Atchison, C., et Gray, J. J. (2009). Host, weather and virological factors drive norovirus epidemiology: Time-series analysis of laboratory surveillance data in England and Wales. PLOS ONE, 4(8), 1-10. https://doi.org/10.1371/journal.pone.0006671 ↩
Ludwig, A., Bigras-Poulin, M., Michel, P., et Bélanger, D. (2010). Risk factors associated with West Nile virus mortality in American Crow populations in southern Quebec. Journal of Wildlife Diseases, 46(1), 195-208. https://doi.org/10.7589/0090-3558-46.1.195 ↩
Ludwig, A., Zheng, H., Vrbova, L., Drebot, M. A., Iranpour, M., et Lindsay, L. R. (2019). Increased risk of endemic mosquito-borne diseases in Canada due to climate change. Canada Communicable Disease Report, 45(4), 90-97. https://doi.org/10.14745/ccdr.v45i04a03 ↩
MacDougall, L., Fyfe, M., Romney, M., Starr, M., et Galanis, E. (2011). Risk factors for Cryptococcus gattii infection, British Columbia, Canada. Emerging Infectious Diseases, 17(2), 193-199. https://doi.org/10.3201/eid1702.101020 ↩
MacDougall, L., Kidd, S. E., Galanis, E., Mak, S., Leslie, M. J., Cieslak, P. R., . . . Bartlett, K. H. (2007). Spread of Cryptococcus gattii in British Columbia, Canada, and detection in the Pacific Northwest, USA. Emerging Infectious Diseases, 13(1), 42-50. https://doi.org/10.3201/eid1301.060827 ↩
Machado-Machado, E. A. (2012). Empirical mapping of suitability to dengue fever in Mexico using species distribution modeling. Applied Geography, 33, 82-93. https://doi.org/10.1016/j.apgeog.2011.06.011 ↩
Maiga, A. W., Deppen, S., Scaffidi, B. K., Baddley, J., Aldrich, M. C., Dittus, R. S., et Grogan, E. L. (2018). Mapping Histoplasma capsulatum exposure, United States. Emerging Infectious Diseases, 24(10), 1835-1839. https://doi.org/10.3201/eid2410.180032 ↩
Mak, S., Morshed, M., et Henry, B. (2010). Ecological niche modeling of Lyme disease in British Columbia, Canada. Journal of Medical Entomology, 47(1), 99-105. https://doi.org/10.1603/033.047.0114 ↩
Maksimovic, Z., Cornwell, M. S., Semren, O., et Rifatbegovic, M. (2017). The apparent role of climate change in a recent anthrax outbreak in cattle. OIE Scientific and Technical Review, 36(3), 959-963. https://doi.org/10.20506/rst.36.3.2727 ↩
Marsden-Haug, N., Goldoft, M., Ralston, C., Limaye, A. P., Chua, J., Hill, H., . . . Chiller, T. (2013). Coccidioidomycosis acquired in Washington state. Clinical Infectious Diseases, 56(6), 847-850. https://doi.org/10.1093/cid/cis1028 ↩
Marston, B. J., Lipman, H. B., et Breiman, R. F. (1994). Surveillance for Legionnaires’ disease: Risk factors for morbidity and mortality. Archives of Internal Medicine, 154(21), 2417-2422. https://doi.org/10.1001/archinte.1994.00420210049006 ↩
Mas-Coma, S., Valero, M. A., et Bargues, M. D. (2008). Effects of climate change on animal and zoonotic helminthiasis. OIE Scientific and Technical Review, 27(2), 443-457. https://doi.org/10.20506/rst.27.2.1822 ↩
McBride, J. A., Gauthier, G. M., et Klein, B. S. (2017). Clinical manifestations and treatment of blastomycosis. Clinics in Chest Medicine, 38(3), 435-449. https://doi.org/10.1016/j.ccm.2017.04.006 ↩
McDonough, E. S., Wisniewski, T. R., Penn, L. A., Chan, D. M., et McNamara, W. J. (1976). Preliminary studies on conidial liberation of Blastomyces dermatitidis and Histoplasma capsulatum. Sabouraudia, 14(2), 199-204. https://doi.org/10.1080/00362177685190271 ↩
McMichael, C., Barnett, J., et McMichael, A. J. (2012). An ill wind? Climate change, migration, and health. Environmental Health Perspectives, 120(5), 646-654. https://doi.org/10.1289/ehp.1104375 ↩
McTaggart, L. R., Brown, E. M., et Richardson, S. E. (2016). Phylogeographic analysis of Blastomyces dermatitidis and Blastomyces gilchristii reveals an association with North American freshwater drainage basins. PLOS ONE, 11(7), 1-22. https://doi.org/10.1371/journal.pone.0159396 ↩
Medlock, J. M., et Vaux, A. G. C. (2011). Assessing the possible implications of wetland expansion and management on mosquitoes in Britain. European Mosquito Bulletin, 29, 38-65. Consulté sur le site : http://e-m-b.org/sites/e-m-b.org/files/EMB%2829%2938-65.pdf ↩
Melles, S. J., Fortin, M. J., Lindsay, K., et Badzinski, D. (2011). Expanding northward: Influence of climate change, forest connectivity, and population processes on a threatened species’ range shift. Global Change Biology, 17(1), 17-31. https://doi.org/10.1111/j.1365-2486.2010.02214.x ↩
Michele, M., Alberto, M., Liana, S., et Francesco, D. (2006). Do environmental factors influence the occurrence of acute meningitis in industrialized countries? An epidemic of varying aetiology in Northern Italy. European Journal of Epidemiology, 21(6), 465-468. https://doi.org/10.1007/s10654-006-9013-1 ↩
Millins, C., Gilbert, L., Medlock, J., Hansford, K., Thompson, D. B. A., et Biek, R. (2017). Effects of conservation management of landscapes and vertebrate communities on Lyme borreliosis risk in the United Kingdom. Philosophical Transactions of the Royal Society B: Biological Sciences, 372(1722), 1-12. https://doi.org/10.1098/rstb.2016.0123 ↩
Minigan, J. N., Hager, H. A., Peregrine, A. S., et Newman, J. A. (2018). Current and potential future distribution of the American dog tick (Dermacentor variabilis, Say) in North America. Ticks and Tick-borne Diseases, 9(2), 354-362. https://doi.org/10.1016/j.ttbdis.2017.11.012 ↩
Mitchell, R., et Kandola, K. (2005). Rabies in the Northwest Territories Part 2: Rabies surveillance in the Northwest Territories. EpiNorth, 17, 4-6.
↩
Monaghan, A. J., Moore, S. M., Sampson, K. M., Beard, C. B., et Eisen, R. J. (2015). Climate change influences on the annual onset of Lyme disease in the United States. Ticks and Tick-borne Diseases, 6(5), 615-622. https://doi.org/10.1016/j.ttbdis.2015.05.005 ↩
Moreno-Madriñán, M. J., et Turell, M. (2018). History of mosquitoborne diseases in the United States and implications for new pathogens. Emerging Infectious Diseases, 24(5), 821-826. https://doi.org/10.3201/eid2405.171609 ↩
Morin, C. W., Semenza, J. C., Trtanj, J. M., Glass, G. E., Boyer, C., et Ebi, K. L. (2018). Unexplored opportunities: Use of climate- and weather-driven early warning systems to reduce the burden of infectious diseases. Current Environmental Health Reports, 5(4), 430-438. https://doi.org/10.1007/s40572-018-0221-0 ↩
Morral-Puigmal, C., Martínez-Solanas, È., Villanueva, C. M., et Basagaña, X. (2018). Weather and gastrointestinal disease in Spain: A retrospective time series regression study. Environment International, 121(1), 649-657. https://doi.org/10.1016/j.envint.2018.10.003 ↩
Morris, S. K., Brophy, J., Richardson, S. E., Summerbell, R., Parkin, P. C., Jamieson, F., . . . Ford-Jones, E. L. (2006). Blastomycosis in Ontario, 1994-2003. Emerging Infectious Diseases, 12(2), 274-279. https://doi.org/10.3201/eid1202.050849 ↩
Mouchtouri, V. A., Verykouki, E., Zamfir, D., Hadjipetris, C., Lewis, H. C., Hadjichristodoulou, C., et The EU SHIPSAN ACT Partnership. (2017). Gastroenteritis outbreaks on cruise ships: Contributing factors and thresholds for early outbreak detection. Eurosurveillance, 22(45), 1-8. https://doi.org/10.2807/1560-7917.ES.2017.22.45.16-00576 ↩
Muchaal, P. K., Parker, S., Meganath, K., Landry, L., et Aramini, J. (2015). Evaluation of a national pharmacy-based syndromic surveillance system. Canada Communicable Disease Report, 41(9), 203-208. https://doi.org/10.14745/ccdr.v41i09a01 ↩
Mukhi, S. N., Andres, E., Demianyk, B., Gammon, B., et Kloeze, H. (2016). KIWI: A technology for public health event monitoring and early warning detection. Online Journal of Public Health Informatics, 8(3), 1-16. https://doi.org/10.5210/ojphi.v8i3.6937 ↩
Murray, D. L. (2003). Relapsing fever. In C. D. Rudolph, A. M. Rudolph, M. K. Hostetter, G. Lister, et N. J. Siegel (Eds.), Rudolph’s pediatrics (21st ed.). New York, NY: McGraw-Hill.
↩
Nakazawa, Y., Williams, R., Peterson, A. T., Mead, P., Staples, E., et Gage, K. L. (2007). Climate change effects on plague and tularemia in the United States. Vector-Borne and Zoonotic Diseases, 7(4), 529-540. https://doi.org/10.1089/vbz.2007.0125 ↩
Nash, D., et Geng, E. (2020). Goal-Aligned, Epidemic Intelligence for the Public Health Response to the COVID-19 Pandemic. American journal of public health, 110(8), 1154–1156. https://doi.org/10.2105/AJPH.2020.305794 ↩
Naughton, D. (2012). The natural history of Canadian mammals. Toronto, ON: University of Toronto Press.
↩
Neil, K., et Berkelman, R. (2008). Increasing incidence of Legionellosis in the United States, 1990–2005: Changing epidemiologic trends. Clinical Infectious Diseases, 47(5), 591-599. https://doi.org/10.1086/590557 ↩
Nelder, M. P., Russell, C. B., Clow, K. M., Johnson, S., Weese, J. S., Cronin, K., . . . Patel, S. N. (2019). Occurrence and distribution of Ambylomma americanum as determined by passive surveillance in Ontario, Canada (1999–2016). Ticks and Tick-borne Diseases, 10(1), 146-155. https://doi.org/10.1016/J.TTBDIS.2018.10.001 ↩
Neumann, G. (2015). H5N1 influenza virulence, pathogenicity and transmissibility: What do we know? Future Virology, 10(8), 971-980. https://doi.org/10.2217/fvl.15.62 ↩
Neumann, G., Noda, T., et Kawaoka, Y. (2009). Emergence and pandemic potential of swine-origin H1N1 influenza virus. Nature, 459(7249), 931-939. https://doi.org/10.1038/nature08157 ↩
Ng, V., et Sargeant, J. M. (2012). A stakeholder-informed approach to the identification of criteria for the prioritization of zoonoses in Canada. PLOS ONE, 7(1), 1-12. https://doi.org/10.1371/journal.pone.0029752 ↩
Ng, V., Fazil, A., Gachon, P., Deuymes, G., Radojević, M., Mascarenhas, M., . . . Ogden, N. H. (2017). Assessment of the probability of autochthonous transmission of chikungunya virus in Canada under recent and projected climate change. Environmental Health Perspectives, 125(6), 1-12. https://doi.org/10.1289/ehp669 ↩
Ng, V., Rees, E. E., Lindsay, R. L., Drebot, M. A., Brownstone, T., Sadeghieh, T., et Khan, S. U. (2019). Could exotic mosquito-borne diseases emerge in Canada with climate change? Canada Communicable Disease Report, 45(4), 98-107. https://doi.org/10.14745/ccdr.v45i04a04 ↩
Nguyen, C., Barker, B. M., Hoover, S., Nix, D. E., Ampel, N. M., Frelinger, J. A., . . . Galgiani, J. N. (2013). Recent advances in our understanding of the environmental, epidemiological, immunological, and clinical dimensions of coccidioidomycosis. Clinical Microbiology Reviews, 26(3), 505-525. https://doi.org/10.1128/CMR.00005-13 ↩
Nicolle, L., Rotstein, C., Bourgault, A., St-Germain, G., Garber, G., et Canadian Infectious Diseases Society Invasive Fungal Registry (1998). Invasive fungal infections in Canada from 1992 to 1994. The Canadian journal of infectious diseases = Journal canadien des maladies infectieuses, 9(6), 347–352. https://doi.org/10.1155/1998/473219 ↩
Nishi, J. S., Stephen, C., et Elkin, B. T. (2002). Implications of agricultural and wildlife policy on management and eradication of bovine tuberculosis and brucellosis in free-ranging wood bison of northern Canada. Annals of the New York Academy of Sciences, 969(1), 236-244. https://doi.org/10.1111/j.1749-6632.2002.tb04385.x ↩
Nyström, M.E., Karltun, J., Keller, C., Andersson Gäre B. (2018). Collaborative and partnership research for improvement of health and social services: researcher’s experiences from 20 projects. Health Research Policy and Systems, 16, 46 https://doi.org/10.1186/s12961-018-0322-0 ↩
O’Brien, S. F., Delage, G., Scalia, V., Lindsay, R., Bernier, F., Dubuc, S., . . . Fearon, M. A. (2016). Seroprevalence of Babesia microti infection in Canadian blood donors. Transfusion, 56(1), 237-243. https://doi.org/10.1111/trf.13339 ↩
O’Connor, S. M., Taylor, C. E., et Hughes, J. M. (2006). Emerging infectious determinants of chronic diseases. Emerging Infectious Diseases, 12(7), 1051-1057. https://doi.org/10.3201/eid1207.060037 ↩
Ogden, N. H. (2014). Lyme disease and climate change. In C. D. Butler (Ed.), Climate change and global health (pp. 145-157). Wallingford, United Kingdom: CABI.
↩
Ogden, N. H. (2017). Climate change and vector-borne diseases of public health significance. FEMS Microbiology Letters, 364(19), 1-8. https://doi.org/10.1093/femsle/fnx186 ↩
Ogden, N. H., AbdelMalik, P., et Pulliam, J. R. C. (2017). Emerging infectious diseases: Prediction and detection. Canada Communicable Disease Report, 43(10), 206-211. https://doi.org/10.14745/ccdr.v43i10a03 ↩
Ogden, N. H., Bigras-Poulin, M., O’Callaghan, C. J., Barker, I. K., Lindsay, L. R., Maarouf, A., . . . Charron, D. (2005). A dynamic population model to investigate effects of climate on geographic range and seasonality of the tick Ixodes scapularis. International Journal for Parasitology, 35(4), 375-389. https://doi.org/10.1016/j.ijpara.2004.12.013 ↩
Ogden, N. H., Bouchard, C., Kurtenbach, K., Margos, G., Lindsay, L. R., Trudel, L., . . . Milord, F. (2010). Active and passive surveillance and phylogenetic analysis of Borrelia burgdorferi elucidate the process of Lyme disease risk emergence in Canada. Environmental Health Perspectives, 118(7), 909-914. https://doi.org/10.1289/ehp.0901766 ↩
Ogden, N. H., et Gachon, P. (2019). Climate change and infectious diseases: What can we expect?. Canada communicable disease report = Releve des maladies transmissibles au Canada, 45(4), 76–80. https://doi.org/10.14745/ccdr.v45i04a01 ↩
Ogden, N. H., et Lindsay, L. R. (2016). Effects of climate and climate change on vectors and vector-borne diseases: Ticks are different. Trends in Parasitology, 32(8), 646-656. https://doi.org/10.1016/j.pt.2016.04.015 ↩
Ogden, N. H., Koffi, J. K., Pelcat, Y., et Lindsay, L. R. (2014c). Environmental risk from Lyme disease in central and eastern Canada: A summary of recent surveillance information. Canada Communicable Disease Report, 40(5), 74-82. https://doi.org/10.14745/ccdr.v40i05a01 ↩
Ogden, N. H., Lindsay, L. R., et Leighton, P. A. (2013). Predicting the rate of invasion of the agent of Lyme disease, Borrelia burgdorferi. Journal of Applied Ecology, 50(2), 510-518. https://doi.org/10.1111/1365-2664.12050 ↩
Ogden, N. H., Lindsay, L. R., et Schofield, S. W. (2015). Methods to prevent tick bites and Lyme disease. Clinics in Laboratory Medicine, 35(4), 883-899. https://doi.org/10.1016/j.cll.2015.07.003 ↩
Ogden, N. H., Lindsay, L. R., Hanincová, K., Barker, I. K., Bigras-Poulin, M., Charron, D. F., . . . Thompson, R. A. (2008). Role of migratory birds in introduction and range expansion of Ixodes scapularis ticks and of Borrelia burgdorferi and Anaplasma phagocytophilum in Canada. Applied and Environmental Microbiology, 74(6), 1780-1790. https://doi.org/10.1128/AEM.01982-07 ↩
Ogden, N. H., Lindsay, L. R., Ludwig, A., Morse, A. P., Zheng, H., et Zhu, H. (2019). Weather-based forecasting of mosquito-borne disease outbreaks in Canada. Canada Communicable Disease Report, 45(5), 127-132. https://doi.org/10.14745/ccdr.v45i05a03 ↩
Ogden, N. H., Margos, G., Artsob, H., et Tsao, J. I. (2014b). Non-rickettsial tick-borne bacteria and the diseases they cause. In D. E. Sonenshine et R. M. Roe (Eds.), Biology of ticks (2nd ed., Vol. 2, pp. 278-312). New York, NY: Oxford University Press.
↩
Ogden, N. H., Pang, G., Ginsberg, H. S., Hickling, G. J., Burke, R. L., Beati, L., et Tsao, J. I. (2018). Evidence for geographic variation in life-cycle processes affecting phenology of the Lyme disease vector Ixodes scapularis (Acari: Ixodidae) in the United States. Journal of Medical Entomology, 55(6), 1386-1401. https://doi.org/10.1093/jme/tjy104 ↩
Ogden, N. H., Radojević, M., Caminade, C., et Gachon, P. (2014a). Recent and projected future climatic suitability of North America for the Asian tiger mosquito Aedes albopictus. Parasites et Vectors, 7, 1-14. https://doi.org/10.1186/s13071-014-0532-4 ↩
Ogden, N. H., Wilson, J. R. U., Richardson, D. M., Hui, C., Davies, S. J., Kumschick, S., . . . Pulliam, J. R. C. (2019). Emerging infectious diseases and biological invasions: A call for a One Health collaboration in science and management. Royal Society Open Science, 6(3), 1-15. https://doi.org/10.1098/rsos.181577 ↩
Ogden, N.H., Maarouf, A., Barker, I.K., Bigras-Poulin, M., Lindsay, L.R., Morshed, M.G., O’Callaghan, C.J., Ramay, F., Waltner-Toews, D. et Charron, D.F. (2006). Climate change and the potential for range expansion of the Lyme disease vector Ixodes scapularis in Canada. International Journal for Parasitology, 36, 63-70.
↩
Oliveira, A. R. S., Piaggio, J., Cohnstaedt, L. W., McVey, D. S., et Cernicchiaro, N. (2018). A quantitative risk assessment (QRA) of the risk of introduction of the Japanese encephalitis virus (JEV) in the United States via infected mosquitoes transported in aircraft and cargo ships. Preventive Veterinary Medicine, 160, 1-9. https://doi.org/10.1016/j.prevetmed.2018.09.020 ↩
O’Lorcain, P. (1995). The effects of freezing on the viability of Toxocara canis and T. cati embryonated eggs. Journal of Helminthology, 69(2), 169-171. https://doi.org/10.1017/S0022149X00014073 ↩
Onozuka, D., et Hashizume, M. (2011). The influence of temperature and humidity on the incidence of hand, foot, and mouth disease in Japan. Science of the Total Environment, 410-411, 119-125. https://doi.org/10.1016/j.scitotenv.2011.09.055 ↩
Ooi, E.-E., et Gubler, D. J. (2009). Global spread of epidemic dengue: the influence of environmental change. Future Virology, 4(6), 571-580. 10.2217/fvl.09.55
↩
O’Reilly, K. M., Auzenbergs, M., Jafari, Y., Liu, Y., Flasche, S., et Lowe, R. (2020). Effective transmission across the globe: the role of climate in COVID-19 mitigation strategies. The Lancet. Planetary health, 4(5), e172. https://doi.org/10.1016/S2542-5196(20)30106-6 ↩
Organisation des Nations unies pour l’alimentation et l’agriculture, et L’organisation mondiale de la santé. (2014). Multicriteria-based ranking for risk management of food-borne parasites: Report of a joint FAO/WHO expert meeting, 3-7 September 2012, FAO headquarters, Rome, Italy (Microbiological risk assessment series 23). Geneva, Switzerland: World Health Organization. Consulté sur le site : https://www.who.int/foodsafety/publications/mra_23/en/ ↩
Organisation mondiale de la santé (OMS). (2019c). Strengthening health security by implementing the International Health Regulations (2005). Consulté sur le site : https://www.who.int/ihr/en/ ↩
Organisation mondiale de la santé. (2013). WHO expert consultation on rabies: Second report (WHO Technical Report Series 982). Geneva, Switzerland: WHO Press. Consulté sur le site : https://apps.who.int/iris/handle/10665/85346 ↩
Organisation mondiale du tourisme et Programme des Nations Unies pour l’environnement. (Eds.). (2008). Climate change and tourism—Responding to global challenges. Madrid, Spain: World Tourism Organization.
↩
Otten A., Fazil, A., Chemeris, A., Breadner, P., et Ng, V. (2019). Prioritization of vector-borne diseases in Canada under current climate and projected climate change.Microbial Risk Analysis, 14. https://doi.org/10.1016/j.mran.2019.100089 ↩
Overy, D. P., McBurney, S., Muckle, A., Lund, L., Lewis, P. J., et Strang, R. (2016). Cryptococcus gattii VGIIb-like variant in white-tailed deer, Nova Scotia, Canada. Emerging Infectious Diseases, 22(6), 1131-1133. https://doi.org/10.3201/eid2206.160081.
↩
Paaijmans, K. P., Cator, L. J., et Thomas, M. B. (2013). Temperature-dependent pre-bloodmeal period and temperature-driven asynchrony between parasite development and mosquito biting rate reduce malaria transmission intensity. PLOS ONE, 8(1), 1-7. https://doi.org/10.1371/journal.pone.0055777 ↩
Paaijmans, K. P., Read, A. F., et Thomas, M. B. (2009). Understanding the link between malaria risk and climate. Proceedings of the National Academy of Sciences of the United States of America, 106(33), 13844-13849. https://doi.org/10.1073/pnas.0903423106 ↩
Pandemic Influenza Outbreak Research Modelling Team. (2009). Modelling an influenza pandemic: a guide for the perplexed. CMAJ, 181(3-4), 171-173. https://doi.org/10.1503/cmaj.090885 ↩
Panic, M., et Ford, J. D. (2013). A review of national-level adaptation planning with regards to the risks posed by climate change on infectious diseases in 14 OECD nations. International Journal of Environmental Research and Public Health, 10(12), 7083-7109. https://doi.org/10.3390/ijerph10127083 ↩
Papp, Z., Clark, R. G., Parmley, E. J., Leighton, F. A., Waldner, C., et Soos, C. (2017). The ecology of avian influenza viruses in wild dabbling ducks (Anas spp.) in Canada. PLOS ONE, 12(5), 1-18. https://doi.org/10.1371/journal.pone.0176297 ↩
Park, Benjamin J., Sigel, K., Vaz, V., Komatsu, K., McRill, C., Phelan, M., . . . Hajjeh, Rana A. (2005). An epidemic of coccidioidomycosis in Arizona associated with climatic changes, 1998-2001. The Journal of Infectious Diseases, 191(11), 1981-1987. https://doi.org/10.1086/430092 ↩
Parkinson, A. J., Evengard, B., Semenza, J. C., Ogden, N., Børresen, M. L., Berner, J., . . . Albihn, A. (2014). Climate change and infectious diseases in the Arctic: Establishment of a circumpolar working group. International Journal of Circumpolar Health, 73(1), 1-7. https://doi.org/10.3402/ijch.v73.25163 ↩
Pastula, D. M., Hoang Johnson, D. K., White, J. L., Dupuis, A. P., Fischer, M., et Staples, J. E. (2015). Jamestown Canyon virus disease in the United States—2000-2013. The American Journal of Tropical Medicine and Hygiene, 93(2), 384-389. https://doi.org/10.4269/ajtmh.15-0196 ↩
Patel, M. M., Pitzer, V. E., Alonso, W. J., Vera, D., Lopman, B., Tate, J., . . . Parashar, U. D. (2013). Global seasonality of rotavirus disease. The Pediatric Infectious Disease Journal, 32(4), e134-e147. https://doi.org/10.1097/INF.0b013e31827d3b68 ↩
Patz, J. A., Githeko, A. K., McCarty, J. P., Hussein, S., Confalonieri, U., et de Wet, N. (2003). Climate change and infectious diseases. In A. J. McMichael, D. H. Campbell-Lendrum, C. F. Corvalán, K. L. Ebi, A. Githeko, J. D. Scheraga, et A. Woodward (Eds.), Climate change and human health: Risks and responses (pp. 103-132). Geneva, Switzerland: World Health Organization.
↩
Patz, J.A., Campbell-Lendrum, D., Holloway, T. et Foley, J.A. (2005). Impact of regional climate change on human health. Nature, 438, 310-7. https://doi.org/10.1038/nature04188.
↩
Paz, S. (2015). Climate change impacts on West Nile virus transmission in a global context. Philosophical Transactions of the Royal Society B: Biological Sciences, 370(1665), 1-11. https://doi.org/10.1098/rstb.2013.0561 ↩
Paz, S., et Semenza, J. C. (2016). El Niño and climate change—Contributing factors in the dispersal of Zika virus in the Americas? The Lancet, 387(10020), 745. https://doi.org/10.1016/S0140-6736(16)00256-7 ↩
Pequeno, P., Mendel, B., Rosa, C., Bosholn, M., Souza, J. L., Baccaro, F., Barbosa, R., et Magnusson, W. (2020). Air transportation, population density and temperature predict the spread of COVID-19 in Brazil. PeerJ, 8, e9322. https://doi.org/10.7717/peerj.9322 ↩
Peters, M., Godfrey, C., McInerney, P., Soares, C. B., Khalil, H., et Parker, D. (2015). The Joanna Briggs Institute reviewers’ manual 2015: Methodology for JBI scoping reviews. Adelaide, Australia: The Joanna Briggs Institute.
↩
Petersen, L. R., Brault, A. C., et Nasci, R. S. (2013a). West Nile virus: Review of the literature. JAMA, 310(3), 308-315. https://doi.org/10.1001/jama.2013.8042 ↩
Petersen, L. R., Carson, P. J., Biggerstaff, B. J., Custer, B., Borchardt, S. M., et Busch, M. P. (2013b). Estimated cumulative incidence of West Nile virus infection in US adults, 1999–2010. Epidemiology et Infection, 141(3), 591-595. https://doi.org/10.1017/s0950268812001070 ↩
Pfister, J. R., Archer, J. R., Hersil, S., Boers, T., Reed, K. D., Meece, J. K., . . . Davis, J. P. (2011). Non-rural point source blastomycosis outbreak near a yard waste collection site. Clinical Medicine et Research, 9(2), 57-65. https://doi.org/10.3121/cmr.2010.958 ↩
Phillips, P., Galanis, E., MacDougall, L., Chong, M. Y., Balshaw, R., Cook, V. J., . . . Thompson, D. (2015). Longitudinal clinical findings and outcome among patients with Cryptococcus gattii infection in British Columbia. Clinical Infectious Diseases, 60(9), 1368-1376. https://doi.org/10.1093/cid/civ041 ↩
Picard-Masson, M., Lajoie, É., Lord, J., Lalancette, C., Marchand, G., Levac, É., . . . Lajoie, L. (2016). Two related occupational cases of Legionella longbeachae infection, Quebec, Canada. Emerging Infectious Diseases, 22(7), 1289-1291. https://doi.org/10.3201/eid2207.160184 ↩
Plummer, F. A., Jones, S. M. (2017), The story of Canada’s Ebola vaccine. Canadian Medical Association Journal, 189 (43), E1326-E1327
↩
Plummer, P. J., McClure, J. T., Menzies, P., Morley, P. S., Van den Brom, R., et Van Metre, D. C. (2018). Management of Coxiella burnetii infection in livestock populations and the associated zoonotic risk: A consensus statement. Journal of Veterinary Internal Medicine, 32(5), 1481-1494. https://doi.org/10.1111/jvim.15229 ↩
Proctor, M. E., Klein, B. S., Jones, J. M., et Davis, J. P. (2002). Cluster of pulmonary blastomycosis in a rural community: Evidence for multiple high-risk environmental foci following a sustained period of diminished precipitation. Mycopathologia, 153(3), 113-120. https://doi.org/10.1023/A:1014515230994 ↩
Public Health Agency of Canada. (2009). Case definitions for communicable diseases under national surveillance. Cananda Communicable Disease Report, 35(Suppl.2).
↩
Raghavan, R. K., Peterson, A. T., Cobos, M. E., Ganta, R., et Foley, D. (2019). Current and future distribution of the lone star tick, Amblyomma americanum (L.) (Acari: Ixodidae) in North America. PLOS ONE, 14(1), 1-13. https://doi.org/10.1371/journal.pone.0209082 ↩
Rajagopalan, S., et Yoshikawa, T. T. (2016). Norovirus infections in long-term care facilities. Journal of the American Geriatrics Society, 64(5), 1097-1103. https://doi.org/10.1111/jgs.14085 ↩
Randazzo, W., Truchado, P., Cuevas-Ferrando, E., Simón, P., Allende, A., et Sánchez, G. (2020). SARS-CoV-2 RNA in wastewater anticipated COVID-19 occurrence in a low prevalence area. Water research, 181, 115942. https://doi.org/10.1016/j.watres.2020.115942 ↩
Randolph, S. E. (2004). Faune, climat et politique: Causes possibles aux récents accroissements des zoonoses à tiques [Fauna, climate and politics: Possible causes for the recent increases in tick-borne zoonoses]. Archives de Pédiatrie, 11(10), 1282-1285. https://doi.org/10.1016/j.arcped.2003.12.019 ↩
Rausch, R. (1956). Studies on the helminth fauna of Alaska: The occurrence of Echinococcus multilocularis Leuckart, 1863, on the mainland of Alaska. The American Journal of Tropical Medicine and Hygiene, 5(6), 1086-1092. https://doi.org/10.4269/ajtmh.1956.5.1086 ↩
Reed, K. D., Meece, J. K., Archer, J. R., et Peterson, A. T. (2008). Ecologic niche modeling of Blastomyces dermatitidis in Wisconsin. PLOS ONE, 3(4), 1-7. https://doi.org/10.1371/journal.pone.0002034 ↩
Reeves, W. C., Hardy, J. L., Reisen, W. K., et Milby, M. M. (1994). Potential effect of global warming on mosquito-borne arboviruses. Journal of Medical Entomology, 31(3), 323-332. https://doi.org/10.1093/jmedent/31.3.323 ↩
Reina Ortiz, M., Le, N. K., Sharma, V., Hoare, I., Quizhpe, E., Teran, E., . . . Izurieta, R. (2017). Post-earthquake Zika virus surge: Disaster and public health threat amid climatic conduciveness. Scientific Reports, 7, 1-10. https://doi.org/10.1038/s41598-017-15706-w ↩
Reisen, W., et Brault, A. C. (2007). West Nile virus in North America: Perspectives on epidemiology and intervention. Pest Management Science, 63(7), 641-646. https://doi.org/10.1002/ps.1325 ↩
Reiter, P. (2001). Climate change and mosquito-borne disease. Environmental Health Perspectives, 109(suppl 1), 141–161.
↩
Reiter, P. (2008). Climate change and mosquito-borne disease: Knowing the horse before hitching the cart. OIE Scientific and Technical Review, 27(2), 383-398. https://doi.org/10.20506/rst.27.2.1804 ↩
Revich, B., Tokarevich, N., et Parkinson, A. J. (2012). Climate change and zoonotic infections in the Russian Arctic. International Journal of Circumpolar Health, 71(1), 1-8. https://doi.org/10.3402/ijch.v71i0.18792 ↩
Rezza, G., Nicoletti, L., Angelini, R., Romi, R., Finarelli, A. C., Panning, M., . . . Cassone, A. (2007). Infection with chikungunya virus in Italy: An outbreak in a temperate region. The Lancet, 370(9602), 1840-1846. https://doi.org/10.1016/s0140-6736(07)61779-6 ↩
Ripoche, M., Campagna, C., Ludwig, A., Ogden, N. H., et Leighton, P. A. (2019). Short-term forecasting of daily abundance of West Nile virus vectors Culex pipiens-restuans (Diptera: Culicidae) and Aedes vexans based on weather conditions in southern Québec (Canada). Journal of Medical Entomology, 56(3), 859-872. https://doi.org/10.1093/jme/tjz002 ↩
Roberts, K. C., Rao, D. P., Bennett, T. L., Loukine, L., et Jayaraman, G. C. (2015). Prevalence and patterns of chronic disease multimorbidity and associated determinants in Canada. Health Promotion and Chronic Disease Prevention Canada, 35(6), 87-94. doi: 10.24095/hpcdp.35.6.01. PMID: 26302227; PMCID: PMC4910465
↩
Rocheleau, J. P., Michel, P., Lindsay, L. R., Drebot, M., Dibernardo, A., Ogden, N. H., . . . Arsenault, J. (2018). Risk factors associated with seropositivity to California serogroup viruses in humans and pet dogs, Quebec, Canada. Epidemiology et Infection, 146(9), 1167-1176. https://doi.org/10.1017/s0950268818001000 ↩
Rodenhouse, N. L., Christenson, L. M., Parry, D., et Green, L. E. (2009). Climate change effects on native fauna of northeastern forests. Canadian Journal of Forest Research, 39(2), 249-263. https://doi.org/10.1139/X08-160 ↩
Roe, C. C., Bowers, J., Oltean, H., DeBess, E., Dufresne, P. J., McBurney, S., . . . Engelthaler, D. M. (2018). Dating the Cryptococcus gattii dispersal to the North American Pacific Northwest. mSphere, 3(1), 1-13. https://doi.org/10.1128/mSphere.00499-17 ↩
Rohr, J. R., Dobson, A. P., Johnson, P. T. J., Kilpatrick, A. M., Paull, S. H., Raffel, T. R., . . . Thomas, M. B. (2011). Frontiers in climate change-disease research. Trends in Ecology et Evolution, 26(6), 270-277. https://doi.org/10.1016/j.tree.2011.03.002 ↩
Romeo-Aznar, V., Paul, R., Telle, O., et Pascual, M. (2018). Mosquito-borne transmission in urban landscapes: The missing link between vector abundance and human density. Proceedings of the Royal Society B: Biological Sciences, 285(1884), 1-9. https://doi.org/10.1098/rspb.2018.0826 ↩
Romero-Lankao, P., Smith, J. B., Davidson, D. J., Diffenbaugh, N. S., Kinney, P. L., Kirshen, P., . . . Villers Ruiz, L. (2014). North America. In V. R. Barros, C. B. Field, D. J. Dokken, M. D. Mastrandrea, K. J. Mach, T. E. Bilir, M. Chatterjee, K. L. Ebi, Y. O. Estrada, R. C. Genova, B. Girma, E. S. Kissel, A. N. Levy, S. MacCracken, P. R. Mastrandrea, et L. L. White (Eds.), Climate change 2014: Impacts, adaptation, and vulnerability. Part B: Regional aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (pp. 1439-1498). Cambridge, England: Cambridge University Press. Consulté sur le site : https://www.ipcc.ch/report/ar5/wg2/ ↩
Rosas, Á. L., et Casadevall, A. (2006). Melanization affects susceptibility of Cryptococcus neoformans to heat and cold. FEMS Microbiology Letters, 153(2), 265-272. https://doi.org/10.1111/j.1574-6968.1997.tb12584.x ↩
Rosenstein, N. E., Perkins, B. A., Stephens, D. S., Popovic, T., et Hughes, J. M. (2001). Meningococcal disease. The New England Journal of Medicine, 344(18), 1378-1388. https://doi.org/10.1056/NEJM200105033441807 ↩
Roussel, M., Pontier, D., Cohen, J.-M., Lina, B., et Fouchet, D. (2016). Quantifying the role of weather on seasonal influenza. BMC Public Health, 16, 1-14. https://doi.org/10.1186/s12889-016-3114-x ↩
Rycroft-Malone, J., Burton, C., Wilkinson, J., Harvey, G., McCormack, B., Baker, R., Dopson, S., Graham, I.D., Staniszewska, S., Thompson, C., Ariss, S., Melville-Richards, L., Williams, L. (2015). Collective action for implementation: a realist evaluation of organisational collaboration in healthcare. Implementation Science, 11(17). https://doi.org/10.1186/s13012-016-0380-z ↩
Safronetz, D., Sloan, A., Stein, D. R., Mendoza, E., Barairo, N., Ranadheera, C., . . . Drebot, M. (2017). Evaluation of 5 commercially available Zika virus immunoassays. Emerging Infectious Diseases, 23(9), 1577-1580. https://doi.org/10.3201/eid2309.162043 ↩
Sage, K. M., Johnson, T. L., Teglas, M. B., Nieto, N. C., et Schwan, T. G. (2017). Ecological niche modeling and distribution of Ornithodoros hermsi associated with tick-borne relapsing fever in western North America. PLOS Neglected Tropical Diseases, 11(10), 1-18. https://doi.org/10.1371/journal.pntd.0006047 ↩
Sagurova I, Ludwig A, Ogden NH, Pelcat Y, Dueymes G, Gachon P. (2019). Predicted northward expansion of the geographic range of the tick vector Amblyomma americanum in North America under future climate conditions. Environmental Health Perspectives, 127(10), 107014.https://doi.org/10.1289/EHP5668 ↩
Sakamoto, R., Ohno, A., Nakahara, T., Satomura, K., Iwanaga, S., Kouyama, Y., . . . Yamaguchi, K. (2009). Legionella pneumophila in rainwater on roads. Emerging Infectious Diseases, 15(8), 1295-1297. https://doi.org/10.3201/eid1508.090317 ↩
Salb, A. L., Barkema, H. W., Elkin, B. T., Thompson, R. C. A., Whiteside, D. P., Black, S. R., . . . Kutz, S. J. (2008). Dogs as sources and sentinels of parasites in humans and wildlife, northern Canada. Emerging Infectious Diseases, 14(1), 60-63. https://doi.org/10.3201/eid1401.071113 ↩
Salkeld DJ, Padgett KA, Jones JH. (2013). A meta-analysis suggesting that the relationship between biodiversity and risk of zoonotic pathogen transmission is idiosyncratic. Ecology Letters, 16(5), 679-686
↩
Samia, N. I., Kausrud, K. L., Heesterbeek, H., Ageyev, V., Begon, M., Chan, K. S., Stenseth, N. C. (2011) Dynamics of the plague-wildlife-human system in Central Asia are controlled by two epidemiological thresholds. Proceedings of the National Academies of Science of the U S A, 108(35), 14527-14532.
↩
Samy, A. M., Elaagip, A. H., Kenawy, M. A., Ayres, C. F., Peterson, A. T., et Soliman, D. E. (2016). Climate change influences on the global potential distribution of the mosquito Culex quinquefasciatus, vector of West Nile virus and lymphatic filariasis. PLOS ONE, 11(10), 1-13. https://doi.org/10.1371/journal.pone.0163863 ↩
Sapp, S. G. H., Rascoe, L. N., Wilkins, P. P., Handali, S., Gray, E. B., Eberhard, M. L., . . . Yabsley, M. J. (2016). Baylisascaris procyonis roundworm seroprevalence among wildlife rehabilitators, United States and Canada, 2012–2015. Emerging Infectious Diseases, 22(12), 2128-2131. https://doi.org/10.3201/eid2212.160467 ↩
Schanzer, D. L., Langley, J. M., et Tam, T. W. S. (2006). Hospitalization attributable to influenza and other viral respiratory illnesses in Canadian children. The Pediatric Infectious Disease Journal, 25(9), 795-800. https://doi.org/10.1097/01.inf.0000232632.86800.8c ↩
Schanzer, D. L., Langley, J. M., et Tam, T. W. S. (2008). Role of influenza and other respiratory viruses in admissions of adults to Canadian hospitals. Influenza and Other Respiratory Viruses, 2(1), 1-8. https://doi.org/10.1111/j.1750-2659.2008.00035.x ↩
Schanzer, D. L., McGeer, A., et Morris, K. (2013a). Statistical estimates of respiratory admissions attributable to seasonal and pandemic influenza for Canada. Influenza and Other Respiratory Viruses, 7(5), 799-808. https://doi.org/10.1111/irv.12011 ↩
Schanzer, D. L., Sevenhuysen, C., Winchester, B., et Mersereau, T. (2013b). Estimating influenza deaths in Canada, 1992-2009. PLOS ONE, 8(11), 1-11. https://doi.org/10.1371/journal.pone.0080481 ↩
Schneeberger, P. M., Wintenberger, C., van der Hoek, W., et Stahl, J. P. (2014). Q fever in the Netherlands 2007-2010: What we learned from the largest outbreak ever. Médecine et Maladies Infectieuses, 44(8), 339-353. https://doi.org/10.1016/j.medmal.2014.02.006 ↩
Schultz-Cherry, S., Olsen, C. W., et Easterday, B. C. (2011). History of swine influenza. Current Topics in Microbiology and Immunology, 370, 21-28. https://doi.org/10.1007/82_2011_197 ↩
Seitz, A. E., Adjemian, J., Steiner, C. A., et Prevots, D. R. (2015). Spatial epidemiology of blastomycosis hospitalizations: Detecting clusters and identifying environmental risk factors. Medical Mycology, 53(5), 447-454. https://doi.org/10.1093/mmy/myv014 ↩
Sejvar, J. J. (2014). Clinical manifestations and outcomes of West Nile virus infection. Viruses, 6(2), 606-623. https://doi.org/10.3390/v6020606 ↩
Semenza, J. C., Lindgren, E., Balkanyi, L., Espinosa, L., Almqvist, M. S., Penttinen, P., et Rocklöv, J. (2016). Determinants and drivers of infectious disease threat events in Europe. Emerging Infectious Diseases, 22(4), 581-589. https://doi.org/10.3201/eid2204 ↩
Septfons, A., Leparc-Goffart, I., Couturier, E., Franke, F., Deniau, J., Balestier, A., . . . De Valk, H. (2016). Travel-associated and autochthonous Zika virus infection in mainland France, 1 January to 15 July 2016. Eurosurveillance, 21(32), 1-7. https://doi.org/10.2807/1560-7917.Es.2016.21.32.30315 ↩
Shafir, S. C., Sorvillo, F. J., Sorvillo, T., et Eberhard, M. L. (2011). Viability of Baylisascaris procyonis eggs. Emerging Infectious Diseases, 17(7), 1293-1295. https://doi.org/10.3201/eid1707.101774 ↩
Shaman, J., Day, J. F., et Stieglitz, M. (2005). Drought-induced amplification and epidemic transmission of West Nile virus in southern Florida. Journal of Medical Entomology, 42(2), 134-141. https://doi.org/10.1093/jmedent/42.2.134 ↩
Shaman, J., Pitzer, V. E., Viboud, C., Grenfell, B. T., et Lipsitch, M. (2010). Absolute humidity and the seasonal onset of influenza in the continental United States. PLOS Biology, 8(2), 1-13. https://doi.org/10.1371/journal.pbio.1000316 ↩
Shapiro, L. L. M., Whitehead, S. A., et Thomas, M. B. (2017). Quantifying the effects of temperature on mosquito and parasite traits that determine the transmission potential of human malaria. PLOS Biology, 15(10), 1-21. https://doi.org/10.1371/journal.pbio.2003489 ↩
Silk, B. J., Foltz, J. L., Ngamsnga, K., Brown, E., Muñoz, M. G., Hampton, L. M., . . . Hicks, L. A. (2013). Legionnaires’ disease case-finding algorithm, attack rates, and risk factors during a residential outbreak among older adults: An environmental and cohort study. BMC Infectious Diseases, 13, 1-8. https://doi.org/10.1186/1471-2334-13-291 ↩
Simmering, J. E., Polgreen, L. A., Hornick, D. B., Sewell, D. K., et Polgreen, P. M. (2017). Weather-dependent risk for Legionnaires’ disease, United States. Emerging Infectious Diseases, 23(11), 1843-1851. https://doi.org/10.3201/eid2311.170137 ↩
Simon, A., Tardy, O., Hurford, A., Lecomte, N., Bélanger, D., et Leighton, P. (2019). Dynamics and persistence of rabies in the Arctic. Polar Research, 38, 1-14.
↩
Simon, J. A., Marrotte, R. R., Desrosiers, N., Fiset, J., Gaitan, J., Gonzalez, A., . . . Millien, V. (2014). Climate change and habitat fragmentation drive the occurrence of Borrelia burgdorferi, the agent of Lyme disease, at the northeastern limit of its distribution. Evolutionary Applications, 7(7), 750-764. https://doi.org/10.1111/eva.12165 ↩
Sivabalan, P., Saboo, A., Yew, J., et Norton, R. (2017). Q fever in an endemic region of North Queensland, Australia: A 10 year review. One Health, 3, 51-55. https://doi.org/10.1016/J.ONEHLT.2017.03.002 ↩
Skog, L., Linde, A., Palmgren, H., Hauska, H., et Elgh, F. (2014). Spatiotemporal characteristics of pandemic influenza. BMC Infectious Diseases, 14, 1-14. https://doi.org/10.1186/1471-2334-14-378 ↩
Skowronski, D. M., Li, Y., Tweed, S. A., Tam, T. W. S., Petric, M., David, S. T., . . . Brunham, R. C. (2007). Protective measures and human antibody response during an avian influenza H7N3 outbreak in poultry in British Columbia, Canada. CMAJ, 176(1), 47-53. https://doi.org/10.1503/cmaj.060204 ↩
Smith, K. R., Woodward, A., Campbell-Lendrum, D., Chadee, D. D., Honda, Y., Liu, Q., . . . Sauerborn, R. (2014). Human health: Impacts, adaptation, and co-benefits. In C. B. Field, V. R. Barros, D. J. Dokken, K. J. Mach, M. D. Mastrandrea, T. E. Bilir, M. Chatterjee, K. L. Ebi, Y. O. Estrada, R. C. Genova, B. Girma, E. S. Kissel, A. N. Levy, S. MacCracken, P. R. Mastrandrea, et L. L. White (Eds.), Climate change 2014: Impacts, adaptation, and vulnerability. Part A: Global and sectoral aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (pp. 709-754). Cambridge, England: Cambridge University Press. Consulté sur le site : https://www.ipcc.ch/report/ar5/wg2/ ↩
Sonenshine, D. E. (2018). Range expansion of tick disease vectors in North America: Implications for spread of tick-borne disease. International Journal of Environmental Research and Public Health, 15(3), 1-9. https://doi.org/10.3390/ijerph15030478 ↩
Souto, A. C. P., Bonfietti, L. X., Ferreira-Paim, K., Trilles, L., Martins, M., Ribeiro-Alves, M., . . . Meyer, W. (2016). Population genetic analysis reveals a high genetic diversity in the Brazilian Cryptococcus gattii VGII population and shifts the global origin from the Amazon rainforest to the semi-arid desert in the northeast of Brazil. PLOS Neglected Tropical Diseases, 10(8), 1-19. https://doi.org/10.1371/journal.pntd.0004885 ↩
Spence, L., Artsob, H., Grant, L., et Th’Ng, C. (1977). St. Louis encephalitis in southern Ontario: Laboratory studies for arboviruses. CMAJ, 116(1), 35-37. Consulté sur le site : https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1879132/ ↩
Stafford, K. C., III., Molaei, G., Little, E. A. H., Paddock, C. D., Karpathy, S. E., et Labonte, A. M. (2018). Distribution and establishment of the lone star tick in Connecticut and implications for range expansion and public health. Journal of Medical Entomology, 55(6), 1561-1568. https://doi.org/10.1093/jme/tjy115 ↩
St-Germain, G., Murray, G., et Duperval, R. (1993). Blastomycosis in Quebec (1981-90): Report of 23 cases and review of published cases from Quebec. Canadian Journal of Infectious Diseases, 4(2), 89-94. https://doi.org/10.1155/1993/249823 ↩
St-Pierre, J., Dufresne, P. J., Carignan, A., Lévesque, É., Bernard, F., Longtin, J., et LeBlanc, L. (2018). Case series: Report of the first two human indigenous cases of Cryptococcus gattii infection in eastern Canada. Mycopathologia, 183(2), 399-406. https://doi.org/10.1007/s11046-017-0215-8 ↩
Sultan, B., Labadi, K., Guégan, J.-F., et Janicot, S. (2005). Climate drives the meningitis epidemics onset in West Africa. PLOS Medicine, 2(1), 0043-0049. https://doi.org/10.1371/journal.pmed.0020006 ↩
Sumi, A., Toyoda, S., Kanou, K., Fujimoto, T., Mise, K., Kohei, Y., . . . Kobayashi, N. (2017). Association between meteorological factors and reported cases of hand, foot, and mouth disease from 2000 to 2015 in Japan. Epidemiology et Infection, 145(14), 2896-2911. https://doi.org/10.1017/S0950268817001820 ↩
Sunenshine, R. H., Anderson, S., Erhart, L., Vossbrink, A., Kelly, P. C., Engelthaler, D., et Komatsu, K. (2007). Public health surveillance for coccidioidomycosis in Arizona. Annals of the New York Academy of Sciences, 1111(1), 96-102. https://doi.org/10.1196/annals.1406.045 ↩
Suresh, S., Forgie, S., et Robinson, J. (2018). Non-polio Enterovirus detection with acute flaccid paralysis: A systematic review. Journal of Medical Virology, 90(1), 3-7. https://doi.org/10.1002/jmv.24933 ↩
Tabachnick, W. J. (2016). Climate change and the arboviruses: Lessons from the evolution of the dengue and yellow fever viruses. Annual Review of Virology, 3, 125-145. https://doi.org/10.1146/annurev-virology-110615-035630 ↩
Takeuchi-Storm, N., Woolsey, I. D., Jensen, P. M., Fredensborg, B. L., Pipper, C. B., et Kapel, C. M. O. (2015). Predictors of Echinococcus multilocularis prevalence in definitive and intermediate hosts: A meta-analysis approach. Journal of Parasitology, 101(3), 297-303. https://doi.org/10.1645/14-645.1 ↩
Tam, B. Y., Martin, I., et Tsuji, L. J. S. (2014). Geospatial analysis between the environment and past incidences of West Nile virus in bird specimens in Ontario, Canada. GeoJournal, 79(6), 805-817. https://doi.org/10.1007/s10708-014-9526-1 ↩
Tamerius, J. D., et Comrie, A. C. (2011). Coccidioidomycosis incidence in Arizona predicted by seasonal precipitation. PLOS ONE, 6(6), 1-7. https://doi.org/10.1371/journal.pone.0021009 ↩
Tang, J. W., Lai, F. Y. L., Nymadawa, P., Deng, Y. M., Ratnamohan, M., Petric, M., . . . Wong, F. Y. W. (2010). Comparison of the incidence of influenza in relation to climate factors during 2000–2007 in five countries. Journal of Medical Virology, 82(11), 1958-1965. https://doi.org/10.1002/jmv.21892 ↩
Tasanee, A., Chayanit, L., et Agsornpien, N. (2015). Does the correlation between rainfall vs dengue hemorrhagic fever incidence always exist? Case study in Phuket, Thailand. Journal of Environmental Research and Development, 9(3), 509-513. Consulté sur le site : http://jerad.org/archiveabstract.php?vol=9etissue=3 ↩
Tatem, A. J., Hay, S. I., et Rogers, D. J. (2006). Global traffic and disease vector dispersal. Proceedings of the National Academy of Sciences of the United States of America, 103(16), 6242-6247. https://doi.org/10.1073/pnas.0508391103 ↩
Tatem, A. J., Huang, Z., Das, A., Qi, Q., Roth, J., et Qiu, Y. (2012). Air travel and vector-borne disease movement. Parasitology, 139(14), 1816-1830. https://doi.org/10.1017/S0031182012000352 ↩
Teixeira, M. de M., Patan, J. S., Taylor, M. L., Gómez, B. L., Theodoro, R. C., de Hoog, S., Engelthaler, D. M., Zancopé-Oliveira, R. M., Felipe, M. S., Barker, B. M. (2016). Worldwide phylogenetic distributions and population dynamics of the genus Histoplasma. PLoS Neglected Tropical Diseases 10, e0004732. https://doi.org/10.1371/journal.pntd.0004732 ↩
Thielman, A. C., et Hunter, F. F. (2007). Photographic key to the adult female mosquitoes (Diptera: Culicidae) of Canada. Canadian Journal of Arthropod Identification, 04. https://doi.org/10.3752/cjai.2007.04 ↩
Thomas-Bachli, A. L., Pearl, D. L., Berke, O., Parmley, E. J., et Barker, I. K. (2015). A comparison of West Nile virus surveillance using survival analyses of dead corvid and mosquito pool data in Ontario, 2002–2008. Preventive Veterinary Medicine, 122(3), 363-370. https://doi.org/10.1016/j.prevetmed.2015.10.007 ↩
Thompson, W. W., Comanor, L., et Shay, D. K. (2006). Epidemiology of seasonal influenza: Use of surveillance data and statistical models to estimate the burden of disease. The Journal of Infectious Diseases, 194(Suppl. 2), S82-S91. https://doi.org/10.1086/507558 ↩
Tinline, R., et Gregory, D. (i2020). Taking the bite out of rabies: The evolution of rabies management in Canada. Toronto, ON: University of Toronto Press.
↩
Tomaselli, M., Kutz, S., Gerlach, C., et Checkley, S. (2017). Local knowledge to enhance wildlife population health surveillance: Conserving muskoxen and caribou in the Canadian Arctic. Biological Conservation, 217, 337-48.
↩
Tomasello, D., et Schlagenhauf, P. (2013). Chikungunya and dengue autochthonous cases in Europe, 2007-2012. Travel Medicine and Infectious Disease, 11(5), 274-284. https://doi.org/10.1016/j.tmaid.2013.07.006 ↩
Tong, D. Q., Wang, J. X. L., Gill, T. E., Lei, H., et Wang, B. (2017). Intensified dust storm activity and valley fever infection in the southwestern United States. Geophysical Research Letters, 44(9), 4304-4312. https://doi.org/10.1002/2017GL073524 ↩
Tossa, P., Deloge-Abarkan, M., Zmirou-Navier, D., Hartemann, P., et Mathieu, L. (2006). Pontiac fever: An operational definition for epidemiological studies. BMC Public Health, 6, 112. https://doi.org/10.1186/1471-2458-6-112 ↩
Towers, S., Chowell, G., Hameed, R., Jastrebski, M., Khan, M., Meeks, J., . . . Harris, G. (2013). Climate change and influenza: The likelihood of early and severe influenza seasons following warmer than average winters. PLOS Currents: Influenza, 1. https://doi.org/10.1371/currents.flu.3679b56a3a5313dc7c043fb944c6f138 ↩
Tricco, A. C., Lillie, E., Zarin, W., O’Brien, K., Colquhoun, H., Kastner, M., . . . Straus, S. E. (2016). A scoping review on the conduct and reporting of scoping reviews. BMC Medical Research Methodology, 16, 1-10. https://doi.org/10.1186/s12874-016-0116-4 ↩
Trudel, L., Veillette, M., Bonifait, L., et Duchaine, C. (2014). Management of the 2012 Legionella crisis in Quebec City: Need for a better communication between resources and knowledge transfer. Frontiers in Microbiology, 5, 1-4. https://doi.org/10.3389/fmicb.2014.00182 ↩
Tsetsarkin, K. A., Vanlandingham, D. L., McGee, C. E., et Higgs, S. (2007). A single mutation in chikungunya virus affects vector specificity and epidemic potential. PLOS Pathogens, 3(12), 1895-1906. https://doi.org/10.1371/journal.ppat.0030201 ↩
Tuite, A. R., Kinlin, L. M., Kuster, S. P., Jamieson, F., Kwong, J. C., McGeer, A., et Fisman, D. N. (2010). Respiratory virus infection and risk of invasive meningococcal disease in central Ontario, Canada. PLOS ONE, 5(11), 1-7. https://doi.org/10.1371/journal.pone.0015493 ↩
Turell, M. J., Britch, S. C., Aldridge, R. L., Kline, D. L., Boohene, C., et Linthicum, K. J. (2013). Potential for mosquitoes (Diptera: Culicidae) from Florida to transmit Rift Valley fever virus. Journal of Medical Entomology, 50(5), 1111-1117. https://doi.org/10.1603/ME13049 ↩
Turell, M. J., Byrd, B. D., et Harrison, B. A. (2013). Potential for populations of Aedes j. japonicus to transmit Rift Valley fever virus in the USA. Journal of the American Mosquito Control Association, 29(2), 133-137. https://doi.org/10.2987/12-6316r.1 ↩
Turell, M. J., Dohm, D. J., et Fonseca, D. M. (2014). Comparison of the potential for different genetic forms in the Culex pipiens complex in North America to transmit Rift Valley fever virus. Journal of the American Mosquito Control Association, 30(4), 253-259. https://doi.org/10.2987/14-6441R.1 ↩
Turell, M. J., Wilson, W. C., et Bennett, K. E. (2010). Potential for North American mosquitoes (Diptera: Culicidae) to transmit Rift Valley fever virus. Journal of Medical Entomology, 47(5), 884-889. https://doi.org/10.1603/ME10007 ↩
Uejio, C. K., Mak, S., Manangan, A., Luber, G., et Bartlett, K. H. (2015). Climatic influences on Cryptococcus gattii populations, Vancouver Island, Canada, 2002-2004. Emerging Infectious Diseases, 21(11), 1989-1996. https://doi.org/10.3201/eid2111.141161 ↩
Valseth, K., Nesbø, C. L., Easterday, W. R., Turner, W. C., Olsen, J. S., Stenseth, N. C., et Haverkamp, T. H. A. (2017). Temporal dynamics in microbial soil communities at anthrax carcass sites. BMC Microbiology, 17, 1-15. https://doi.org/10.1186/s12866-017-1111-6 ↩
van Leuken, J. P. G., Swart, A. N., Droogers, P., van Pul, A., Heederik, D., et Havelaar, A. H. (2016). Climate change effects on airborne pathogenic bioaerosol concentrations: A scenario analysis. Aerobiologia, 32(4), 607-617. https://doi.org/10.1007/s10453-016-9435-5 ↩
Vander Kelen, P., Downs, J. A., Unnasch, T., et Stark, L. (2014). A risk index model for predicting eastern equine encephalitis virus transmission to horses in Florida. Applied Geography, 48, 79-86. https://doi.org/10.1016/j.apgeog.2014.01.012 ↩
Varia, M., Wilson, S., Sarwal, S., McGeer, A., Gournis, E., Galanis, E., et Henry, B. (2003). Investigation of a nosocomial outbreak of severe acute respiratory syndrome (SARS) in Toronto, Canada. CMAJ, 169(4), 285-292. Consulté sur le site : http://www.cmaj.ca/content/169/4/285.long ↩
Varrin, R., Bowman, J., et Gray, P. A. (2007). The known and potential impacts of climate change on biodiversity in Ontario’s terrestrial ecosystems: Case studies and recommendations for adaptation. (Climate Change Research Report CCRR-09). Sault Ste. Marie, ON: Ontario Ministry of Natural Resources Consulté sur le site : http://www.climateontario.ca/MNR_Publications/276927.pdf ↩
Venturi, G., Di Luca, M., Fortuna, C., Remoli, M. E., Riccardo, F., Severini, F., . . . Rizzo, C. (2017). Detection of a chikungunya outbreak in Central Italy, August to September 2017. Eurosurveillance, 22(39), 1-4. https://doi.org/10.2807/1560-7917.ES.2017.22.39.17-00646 ↩
Viboud, C., Pakdaman, K., Boëlle, P.-Y., Wilson, M. L., Myers, M. F., Valleron, A.-J., et Flahault, A. (2004). Association of influenza epidemics with global climate variability. European Journal of Epidemiology, 19(11), 1055-1059. https://doi.org/10.1007/s10654-004-2450-9 ↩
Villeneuve, A., Polley, L., Jenkins, E., Schurer, J., Gilleard, J., Kutz, S., . . . Gagné, F. (2015). Parasite prevalence in fecal samples from shelter dogs and cats across the Canadian provinces. Parasites et Vectors, 8, 1-10. https://doi.org/10.1186/s13071-015-0870-x ↩
Vugia, D. J., Wheeler, C., Cummings, K. C., et Karon, A. (2009). Increase in coccidioidomycosis—California, 2000-2007. MMWR Morbidity and Mortality Weekly Report, 58(5), 105-109. Consulté sur le site : https://www.cdc.gov/mmwr/preview/mmwrhtml/mm5805a1.htm ↩
Walsh, M. G., de Smalen, A. W., et Mor, S. M. (2018). Climatic influence on anthrax suitability in warming northern latitudes. Scientific Reports, 8, 1-9. https://doi.org/10.1038/s41598-018-27604-w ↩
Wang, C., Liu, L., Hao, X., Guo, H., Wang, Q., Huang, J., He, N., Yu, H., Lin, X., Pan, A., Wei, S., Wu, T. (2020). Evolving Epidemiology and Impact of Non-pharmaceutical Interventions on the Outbreak of Coronavirus Disease 2019 in Wuhan, China. JAMA.https://doi.org/10.1101/2020.03.03.20030593 ↩
Wang, G., Minnis, R. B., Belant, J. L., et Wax, C. L. (2010). Dry weather induces outbreaks of human West Nile virus infections. BMC Infectious Diseases, 10, 1-7. https://doi.org/10.1186/1471-2334-10-38 ↩
Wang, J., Ogden, N. H., et Zhu, H. (2011). The impact of weather conditions on Culex pipiens and Culex restuans (Diptera: Culicidae) abundance: A case study in Peel Region. Journal of Medical Entomology, 48(2), 468-475. https://doi.org/10.1603/ME10117 ↩
Wang, P., Goggins, W. B., et Chan, E. Y. Y. (2016). Hand, foot and mouth disease in Hong Kong: A time-series analysis on its relationship with weather. PLOS ONE, 11(8), 1-12. https://doi.org/10.1371/journal.pone.0161006 ↩
Warren, F. J., et Lemmen, D. S. (éd.) (2014). Vivre avec les changements climatiques au Canada : perspectives des secteurs relatives aux impacts et à l’adaptation. Ottawa, ON: Gouvernement du Canada. Consulté sur le site https://publications.gc.ca/site/fra/9.643395/publication.html ↩
Webster, D., Dimitrova, K., Holloway, K., Makowski, K., Safronetz, D., et Drebot, M. A. (2017). California serogroup virus infection associated with encephalitis and cognitive decline, Canada, 2015. Emerging Infectious Diseases, 23(8), 1423-1424. https://doi.org/10.3201/eid2308.170239 ↩
Weissenböck, H., Kolodziejek, J., Url, A., Lussy, H., Rebel-Bauder, B., et Nowotny, N. (2002). Emergence of Usutu virus, an African mosquito-borne Flavivirus of the Japanese encephalitis virus group, central Europe. Emerging Infectious Diseases, 8(7), 652-656. https://doi.org/10.3201/eid0807.020094 ↩
White, A. N. J., Ng, V., Spain, C. V., Johnson, C. C., Kinlin, L. M., et Fisman, D. N. (2009). Let the sun shine in: Effects of ultraviolet radiation on invasive pneumococcal disease risk in Philadelphia, Pennsylvania. BMC Infectious Diseases, 9. https://doi.org/10.1186/1471-2334-9-196 ↩
Williams, C. R., Mincham, G., Faddy, H., Viennet, E., Ritchie, S. A., et Harley, D. (2016). Projections of increased and decreased dengue incidence under climate change. Epidemiology et Infection, 144(14), 3091-3100. https://doi.org/10.1017/s095026881600162x ↩
Wilson, W. C., Kim, I. J., Trujillo, J. D., Sunwoo, S. Y., Noronha, L. E., Urbaniak, K., . . . Richt, J. A. (2018). Susceptibility of white-tailed deer to Rift Valley fever virus. Emerging Infectious Diseases, 24(9), 1717-1719. https://doi.org/10.3201/eid2409.180265 ↩
Wong, S.S.Y., Yip, C.C.Y., Lau, S.K.P. et Yuen, K.Y. (2010). Human enterovirus 71 and hand, foot and mouth disease. Epidemiology et Infection, 13, 1071-89. https://doi.org/10.1017/S0950268809991555.
↩
Wood, D. M., Dang, P. T., et Ellis, R. A. (1979). Part 6 The mosquitoes of Canada—Diptera: Culicidae. In The insects and arachnids of Canada. Ottawa, ON: Government of Canada. Consulté sur le site : http://publications.gc.ca/pub?id=9.811351etsl=0 ↩
Wormser, G. P., Dattwyler, R. J., Shapiro, E. D., Halperin, J. J., Steere, A. C., Klempner, M. S., . . . Nadelman, R. B. (2006). The clinical assessment, treatment, and prevention of Lyme disease, human granulocytic anaplasmosis, and babesiosis: Clinical practice guidelines by the Infectious Diseases Society of America. Clinical Infectious Diseases, 43(9), 1089-1134. https://doi.org/10.1086/508667 ↩
Wu, J. T., Leung, K., et Leung, G. M. (2020). Nowcasting and forecasting the potential domestic and international spread of the 2019-nCoV outbreak originating in Wuhan, China: A modelling study. The Lancet,395(10225), 689-697. https://doi.org/10.1016/S0140-6736(20)30260-9 ↩
Xiao, F.-Z., Zhang, Y., Deng, Y.-Q., He, S., Xie, H.-G., Zhou, X.-N., et Yan, Y.-S. (2014). The effect of temperature on the extrinsic incubation period and infection rate of dengue virus serotype 2 infection in Aedes albopictus. Archives of Virology, 159(11), 3053-3057. https://doi.org/10.1007/s00705-014-2051-1 ↩
Yang, H. M., Macoris, M. L. G., Galvani, K. C., Andrighetti, M. T. M., et Wanderley, D. M. V. (2009). Assessing the effects of temperature on the population of Aedes aegypti, the vector of dengue. Epidemiology et Infection, 137(8), 1188-1202. https://doi.org/10.1017/s0950268809002040 ↩
Yansouni, C.P., Pernica, J.M., et Goldfarb, D. (2016). Enteric parasites in Arctic communities: Tip of the iceberg? Trends in Parasitology, 32(11), 834-38.
↩
Yoo, E. H., Chen, D., Diao, C., et Russell, C. (2016). The effects of weather and environmental factors on West Nile virus mosquito abundance in Greater Toronto Area. Earth Interactions, 20(3), 1-22. https://doi.org/10.1175/ei-d-15-0003.1 ↩
Yu, G., Li, Y., Cai, J., Yu, D., Tang, J., Zhai, W., . . . Qin, J. (2019). Short-term effects of meteorological factors and air pollution on childhood hand-foot-mouth disease in Guilin, China. Science of the Total Environment, 646, 460-470. https://doi.org/10.1016/j.scitotenv.2018.07.329 ↩
Yue, R. P. H., et Lee, H. F. (2018). Pre-industrial plague transmission is mediated by the synergistic effect of temperature and aridity index. BMC Infectious Diseases, 18(1), 134. https://doi.org/10.1186/s12879-018-3045-5 ↩
Zhang, T., Wu, Q., et Zhang, Z. (2020). Probable Pangolin Origin of SARS-CoV-2 Associated with the COVID-19 Outbreak. Current biology : CB, 30(8), 1578. https://doi.org/10.1016/j.cub.2020.03.063 ↩
Zhao, D., Wang, L., Cheng, J., Xu, J., Xu, Z., Xie, M., . . . Su, H. (2017). Impact of weather factors on hand, foot and mouth disease, and its role in short-term incidence trend forecast in Huainan City, Anhui province. International Journal of Biometeorology, 61(3), 453-461. https://doi.org/10.1007/s00484-016-1225-9 ↩
Zhao, Q., Li, S., Cao, W., Liu, D.-L., Qian, Q., Ren, H., . . . Guo, Y. (2018). Modeling the present and future incidence of pediatric hand, foot, and mouth disease associated with ambient temperature in mainland China. Environmental Health Perspectives, 126(4), 1-11. https://doi.org/10.1289/EHP3062 ↩
Zheng, H., Drebot, M. A., et Coulthart, M. B. (2014). West Nile virus in Canada: Ever-changing, but here to stay. Canada Communicable Disease Report, 40(10), 173-177. https://doi.org/10.14745/ccdr.v40i10a01 ↩
Zhu, F., Xu, W., Xia, J., Liang, Z., Liu, Y., Zhang, X., . . . Wang, N. (2014). Efficacy, safety, and immunogenicity of an enterovirus 71 vaccine in China. The New England Journal of Medicine, 370(9), 818-828. https://doi.org/10.1056/NEJMoa1304923 ↩