Ogden, N. H., Bouchard, C., Brankston, G., Brown, E. M., Corrin, T., Dibernardo, A., Drebot, M. A., Fisman, D. N., Galanis, E., Greer, A., Jenkins, E., Kus, J. V., Leighton, P. A., Lindsay, L. R., Lowe, A.-M., Ludwig, A., Morris, S. K., Ng, V., Vrbova, L., Waddell, L., & Wood, H. (2022). Infectious Diseases. In P. Berry & R. Schnitter (Eds.), Health of Canadians in a Changing Climate: Advancing our Knowledge for Action. Ottawa, ON: Government of Canada.
Lead Author
Nick H. Ogden (Public Health Agency of Canada)
Contributing Authors
Catherine Bouchard (Public Health Agency of Canada)
Gabrielle Brankston (University of Guelph)
Elizabeth M. Brown (Public Health Ontario)
Tricia Corrin (Public Health Agency of Canada)
Antonia Dibernardo (Public Health Agency of Canada)
Michael A. Drebot (Public Health Agency of Canada)
David N. Fisman (University of Toronto)
Eleni Galanis (BC Centre for Disease Control and University of British Columbia)
Amy Greer (University of Guelph)
Emily Jenkins (University of Saskatchewan)
Julianne V. Kus (Public Health Ontario and University of Toronto)
Patrick A. Leighton (Université de Montréal)
Robbin Lindsay (Public Health Agency of Canada)
Anne-Marie Lowe (Public Health Agency of Canada)
Antoinette Ludwig (Public Health Agency of Canada)
Shaun K. Morris (Hospital for Sick Children and University of Toronto)
Climate change is affecting the risk from infectious diseases. There is evidence that the recent emergence of Lyme disease in Canada has been driven by climate warming, making more of Canada suitable for the ticks that carry the disease. Emergence of other insect-borne diseases, such as eastern equine encephalitis, could have been facilitated by a warming climate, and epidemics of West Nile virus infection have likely been driven by variability in weather and climate, which will increase with climate change. The risk from a very wide range of other infectious diseases is also known to be sensitive to weather and climate. Changes to geographic and seasonal patterns of these diseases in North America, and increased risk of importation of climate-sensitive diseases from further afield, are likely to pose increased risks to Canadians in coming decades. Adaptation measures include assessments of risk and vulnerability, integrated surveillance and early warning systems using emerging technologies, and a “One Health” approach that integrates human, animal, and environmental health.
Key Messages
Under climate change, many diseases considered “climate-sensitive” are more likely to emerge or re-emerge globally and in Canada. These diseases include those transmitted by arthropod vectors (such as West Nile virus, Lyme disease), those directly transmitted from animals (zoonoses such as rabies, hantavirus pulmonary syndrome), those directly transmitted human-to-human (such as seasonal influenza, enterovirus infections), and those that can be acquired by inhalation from environmental sources (such as Cryptococcus infection, Legionnaires’ disease).
Infectious diseases new to Canada may spread northward from the United States, and from elsewhere in the world, carried by people and goods, or by wild animals. The indirect socio-economic effects of climate change may affect the capacity of nations to prevent and control infectious diseases globally, increasing the likelihood that new diseases will come into Canada through human travel and migration.
Climate change is expected to make the Canadian environment more suitable for arthropod vectors (such as mosquitoes and ticks) and transmission of new infectious diseases. For example, mosquito-borne diseases already in Canada such as West Nile virus, which usually cause a limited number of infections each year, may produce epidemics under a more variable climate with more frequent extreme weather events.
Potential effects of climate change on infectious diseases are identified by modelling studies, while disease surveillance has identified changes in occurrence of infectious diseases, and in some cases linked these changes to recent effects of climate change. These studies are largely restricted to diseases that humans acquire from arthropod vectors (insects and ticks) and directly from animals.
Canada has high adaptive capacity to cope with infectious diseases given its robust national public health surveillance and response tied into national and international networks, a strong health system, and capacity for technological innovations. Canada is also a leader in “One Health” approaches that consider human, animal, and environmental factors together, using knowledge from many disciplines and sectors. Such approaches are essential to planning for emerging and re-emerging infectious diseases, including those related to climate change.
Canada is also increasing its capacity to respond to effects of climate change on infectious diseases. This capacity will be enhanced by big data and modern genomic technologies, Earth observation from satellites, web crawling, and “citizen science” approaches to surveillance for climate change impacts on infectious diseases.
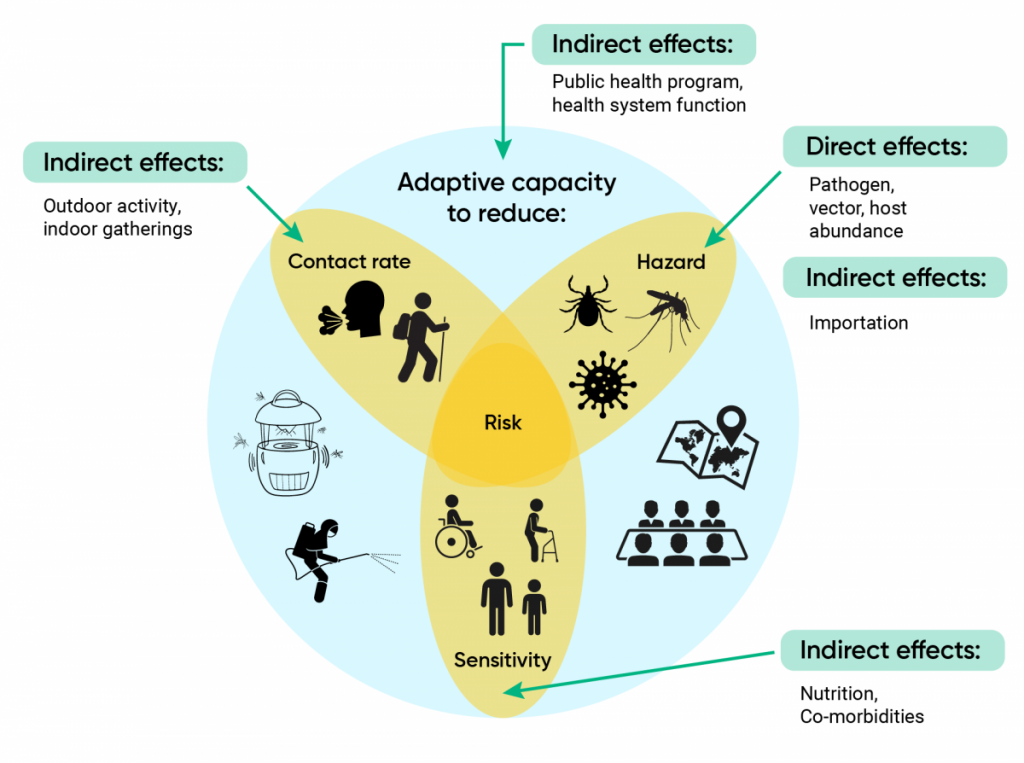
Components of vulnerability to infectious diseases in the context of climate change.
Figure 6.1
The three intersecting components of risk are hazard, contact rate (which, with hazard, determines exposure), and sensitivity. Adaptation (represented by the blue background disc) depends on the capacity to minimize, and respond to changes in each of these three components of risk. Green arrows show direct and indirect effects of climate change.
Overview of the Impacts of Climate Change on Infectious Diseases
Health Impact or Hazard Category
Climate-Related Causes
Possible Health Effects
Infectious diseases transmitted by arthropod vectors
Faster reproduction rates and greater survival, leading to increased abundance and geographic range of vectors found in Canada
Effects of weather variability and extreme weather events on reproduction rates and survival of mosquitoes that lead to rapid changes in populations
Faster reproduction rates and greater survival of exotic vectors once they are carried into Canada, making it more likely that exotic vectors and the diseases they carry (pathogens) can become established
Faster development of pathogens in mosquito vectors
Increased incidence of Canada-endemic vector-borne diseases (e.g., Lyme disease, West Nile virus infection, eastern equine encephalitis)
Increased epidemics of Canada-endemic mosquito-borne diseases (West Nile virus infection, eastern equine encephalitis, California serogroup viruses)
Spread of US-endemic tick-borne (e.g., monocytic ehrlichiosis) and mosquito-borne (e.g., La Crosse virus infection ) diseases into Canada
Increasing risk of autochthonous transmission of tropical/subtropical Aedes mosquito-borne diseases (dengue, chikungunya, Zika)
Impacts on health services
Infectious diseases directly transmitted by animals (zoonotic diseases)
Changes to rates of reproduction and survival of wild animal reservoir hosts and other species resulting in changes in geographic ranges and levels of hazard for, and contact rates with, humans
Effects of weather on reproduction rates and survival of species such as rodents that can lead to rapid changes in their abundance
Changes (increases in some locations, decreases in others) in geographic and temporal patterns of risk of directly transmitted zoonotic diseases (such as rabies, brucellosis)
Possible increasing frequency of outbreaks of some rodent-borne diseases (such as hantavirus pulmonary syndrome)
Impacts on health services
Infectious diseases acquired by inhalation from environmental sources
Possible increased survival and reproduction of fungi in soils and other land-based environments
Possible increase in multiplication of Legionella bacteria due to increased use of air conditioning
Increased incidence, and changing geographic ranges of air-borne and aerosolized fungal infections (such as cryptococcosis)
Increased outbreaks of legionnellosis
Impacts on health services
Emerging infectious diseases
Climate-change induced changes to the ecology of zoonotic diseases globally increases the possibility of emergence, spillover into humans and spread to Canada
Increased international travel, including human population migration, enhances global spread of zoonotic diseases that are also capable of human-to-human transmission
Increased likelihood and frequency of epidemics and pandemics (such as SARS and COVID-19) including water-borne, food-borne, vector-borne, and zoonotic infectious diseases
Increased transmission of pathogens and infectious diseases, potentially leading to not only physical health impacts, but also impacts on mental, spiritual, and psychological health
Impacts on health services
List of Acronyms
CSGV California serogroup viruses
CVV Cache Valley virus
EEEV eastern equine encephalitis virus
EIP extrinsic incubation period
GCM global climate models
GHG greenhouse gas
GOARN Global Outbreak Alert and Response Network
GPHIN Global Public Health Intelligence Network
HFMD hand-foot-and-mouth disease
IHR International Health Regulations
JCV Jamestown Canyon virus
JE Japanese encephalitis
LACV La Crosse encephalitis virus
MCDA multi-criteria decision analysis
RCP representative concentration pathways
RMSF Rocky Mountain spotted fever
RRA rapid risk assessment
RVFV Rift Valley fever virus
SINV Sindbis virus
SLEV St. Louis encephalitis virus
SSHV Snowshoe Hare virus
USUV Usutu virus
VEE Venezuelan equine encephalitis
WGS whole genome sequencing
WHO World Health Organization
WNV West Nile virus
YF yellow fever
6.1
Introduction
Share
Infectious diseases continue to emerge and re-emerge globally, illustrated by epidemics and pandemics of new and existing diseases, such as coronavirus disease 2019 (COVID-19) caused by the SARS-CoV-2 coronavirus, Ebola in Africa, Middle East respiratory syndrome coronavirus infection in the Middle East, and Zika, chikungunya, yellow fever, and dengue virus infections in the Americas. Many infectious diseases are climate-sensitive; as a result, transmission of these diseases may be affected directly or indirectly by climate change. In Canada, it is expected that current infectious disease risks will increase and new diseases will emerge as the climate warms, causing concern for human health. This chapter considers the impacts of climate change on risks from infectious diseases of importance for public health in Canada. It expands on previous national climate change and health assessments to identify the state of knowledge of current and projected risks from infectious diseases, assesses vulnerability, and identifies where public health efforts may be required to protect the Canadian population. Infectious diseases related to the impacts of climate change on pathogen transmission in water and food are discussed in Chapter 7: Water Quality, Quantity, and Security, and Chapter 8: Food Safety and Security, respectively.
The demographic makeup of Canadians is changing, with an aging population (Statistics Canada, 2016), and more people affected by chronic illnesses, both of which can change the sensitivity of people to infectious disease hazards (Dye, 2014). In this chapter, risk is considered to have two main components: likelihood of exposure and sensitivity (i.e., severity of outcome), while the vulnerability of the population is considered to be the risk combined with the adaptive capacity (Figure 6.1). This is similar to other versions in the literature of the relationship between risk and vulnerability, where vulnerability is a component of risk that combines sensitivity, exposure, and adaptive capacity of the human population (IPCC, 2014). Exposure likelihood depends on the level of hazard, which is the number of infective organisms (i.e., infective humans, micro-organisms, arthropod vectors, animal reservoir hosts) in an individual’s environment, and the rate of contact of uninfected humans with the hazard (Figure 6.1). In the context of infectious diseases, adaptive capacity is the capacity of public health systems to identify, prevent, and control disease, and of health systems to minimize the impact of disease through rapid and effective treatment. Climate change may affect each of the components of vulnerability by direct effects on the existence and level of hazard, by indirect effects on rates of contact with the hazard, by increasing or decreasing population sensitivity, and by affecting adaptive capacity itself (Figure 6.1).
Components of vulnerability to infectious diseases in the context of climate change.
Figure 6.1
The three intersecting components of risk are hazard, contact rate (which, with hazard, determines exposure), and sensitivity. Adaptation (represented by the blue background disc) depends on the capacity to minimize, and respond to changes in each of these three components of risk. Green arrows show direct and indirect effects of climate change.
6.1.1
Infectious Disease Emergence and Re-Emergence
Share
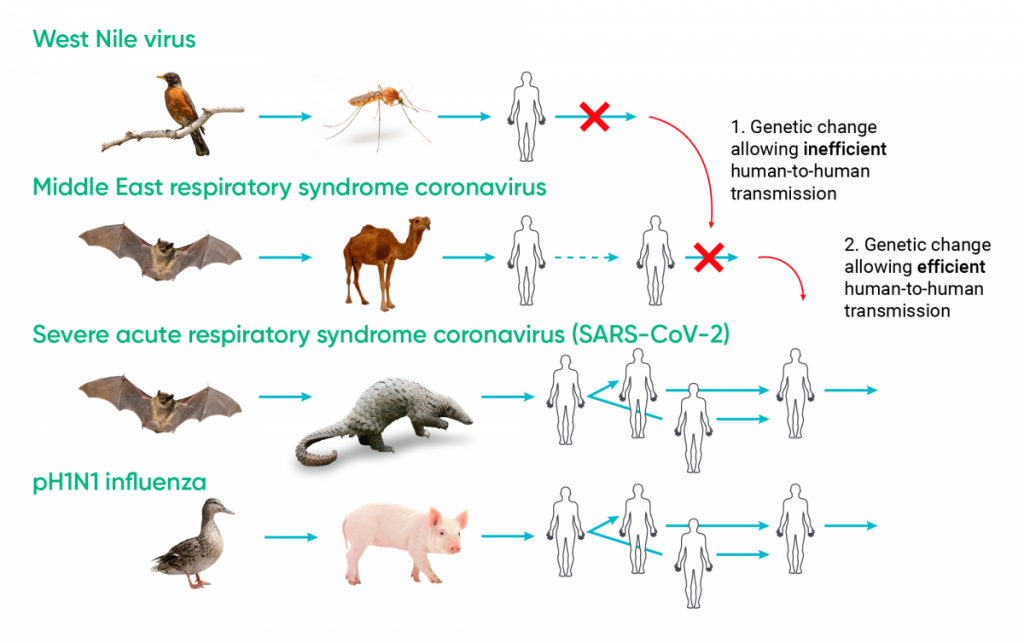
Infectious diseases emerge through changes in their geographic ranges, “spillover,” and “adaptive emergence.” Spillover is when some environmental or socio-economic change allows an animal pathogen already transmissible to humans to come into contact with humans (e.g., Nipah virus). Adaptive emergence is due to genetic change in a micro-organism infecting animals, usually wildlife, that results in it becoming transmissible to humans from the animal (i.e., it becomes a zoonosis) and perhaps transmissible among humans (e.g., SARS-CoV) (Ogden et al., 2017).
There are multiple drivers of disease emergence, which include those associated with globalization and environment. These include climate change, social and demographic changes, and changes in public health systems and policies (Semenza et al., 2016). Endemic diseases can re-emerge (i.e., increase in incidence, or resurge as epidemics) through the same drivers. Climate change may directly affect infectious disease emergence and re-emergence through effects on the survival of pathogens, on survival and reproduction rates of arthropod vectors (e.g., mosquitos, ticks, and fleas), and, in the case of zoonoses, on the abundance of animal reservoir hosts. These are factors that determine the potential for a pathogen to propagate among humans or, for zoonoses, animal hosts, which is described by the basic reproduction number R0. If R0 is one or higher (in which case one infection results in at least one more infection), the pathogen may persist and spread, but if R0 is less than one, it will die out (Anderson & May, 1991). The direct effects of climate change that cause disease emergence in a particular location are effectively those that change R0 from less than one to one or higher in that location.
Climate change may have indirect impacts on disease emergence and re-emergence by affecting other environmental and social changes, and by reducing the ability of public health systems to respond (e.g., extreme weather events may disrupt public health capacity to control disease outbreaks). The effects of climate change on ecosystems, including on biodiversity, may alter the hazards posed by zoonoses through complex effects on wildlife communities (Altizer et al., 2013; Cable et al., 2017). Other changes that can affect disease emergence and re-emergence may be related to public health adaptation initiatives to reduce broader health risks of climate change. For example, efforts to reduce heat islands in urban areas through the greening of cities (Beaudoin & Gosselin, 2016) and actions to manage floods (see Chapter 3: Natural Hazards) may increase zoonosis hazards from wildlife and vector-borne diseases (Medlock & Vaux, 2011; Millins et al., 2017). Increased use of air conditioning to combat urban heat could increase risks of legionellosis (Fitzhenry et al., 2017).
Climate change may have negative impacts on economies, particularly those of low- and middle-income countries, which could directly, or indirectly through an increase in conflicts, reduce infectious disease control and contribute to increasing densities of infectious agents (Ogden, 2017). Economic impacts and dislocation may simultaneously drive increased economic or refugee migration, increasing importation of infectious diseases to Canada from abroad (Ogden, 2017). In addition, if health systems are not climate-resilient (e.g., resilient to outages of power and communication systems associated with extreme weather events) (see Chapter 10: Adaptation and Health System Resilience), impacts may reduce the capacity to detect and respond to emerging or re-emerging infectious diseases (Mayhew & Hanefeld, 2014; Ebi et al., 2018; Global Commission on Adaptation, 2019). The range of projected climate change includes long-term changes in temperature and precipitation patterns, increased climate variability, and increased frequency of extreme weather events, which will vary among geographic regions in Canada (Bush & Lemmen, 2019). These changes will directly and indirectly affect different infectious disease risks idiosyncratically (Ogden & Lindsay, 2016).
6.1.2
Infectious Diseases in Previous Climate Change and Health Assessments
Share
Since 2008, there have been six national, regional, or international assessments on the effects of climate change on infectious disease risks and vulnerability (Table 6.1). There were two assessments in Canada — one that focused on health in 2008 (Charron et al., 2008) and one that included health in a broader Government of Canada report on climate change impacts and adaptation in 2014 (Berry et al., 2014a). International assessments relevant to Canada included chapters on health and on North America from the Intergovernmental Panel on Climate Change (IPCC) (Romero-Lankao et al., 2014; Smith et al., 2014) and national assessments in the United States (Beard et al., 2016; Ebi et al., 2018).
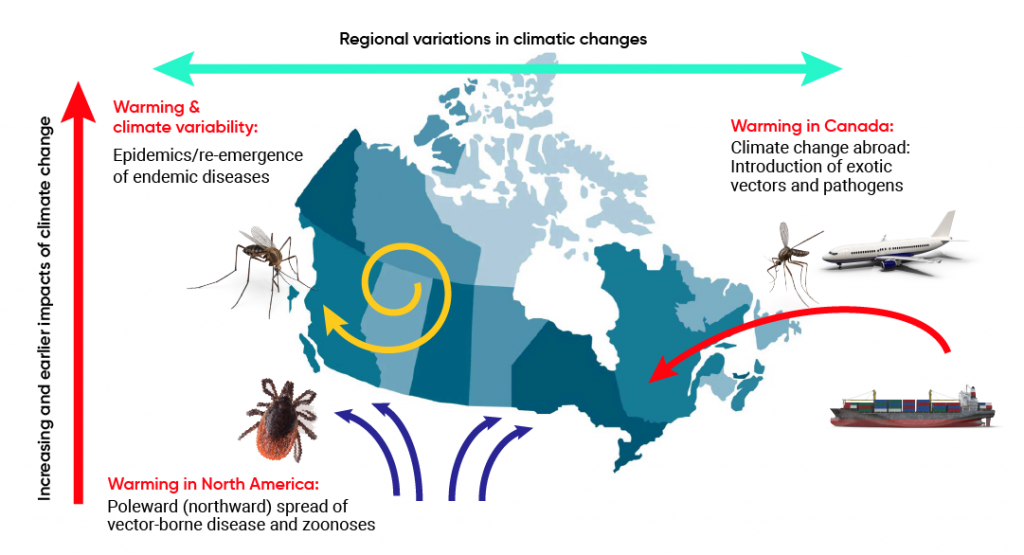
All assessments identified arthropod-borne diseases (i.e., those transmitted by arthropods such as mosquitoes, ticks, and fleas) as most climate-sensitive, with climate determining the occurrence and abundance via effects on vector survival and reproduction, and on pathogen development in some vectors. The assessments suggest that climate warming is likely to increase risks from these diseases, but most assessments indicated moderate confidence in identifying which vector-borne diseases will be affected and the magnitude of the effects. This is due to the complexity of vector-borne disease transmission cycles. Multiple non-climatic determinants, including environmental and land-use changes, particularly for vector-borne zoonoses such as West Nile virus (WNV) and Lyme disease, and control efforts, particularly for exotic mosquito-borne diseases such as malaria and dengue, are important for determining the occurrence and abundance of vectors and pathogens. Previous assessments highlighted the climatic sensitivity of endemic hazard sensitivity in Canada and the United States, including plague and hantavirus infections that have a rodent reservoir; Lyme disease and other tick-borne diseases, such as Rocky Mountain spotted fever (RMSF); and endemic mosquito-borne diseases caused by WNV, eastern equine encephalitis virus (EEEV), and California serogroup viruses. The assessments identified that changes to geographic ranges and length of transmission seasons are particularly likely under climate change (Charron et al., 2008; Berry et al., 2014a; Smith et al., 2014; Beard et al., 2016; Ebi et al., 2018). Figure 6.2 illustrates the pathways through which climate change can affect infectious disease risks in Canada, according to previous assessments.
Climate change effects on infectious disease risks in Canada.
Source
Ogden & Gachon, 2019.
The 2008 Canadian assessment provided model-based projections of the northward spread of Lyme disease from the United States into Canada associated with climate change–driven range expansion of the tick vector Ixodes scapularis (Charron et al., 2008). Subsequently, the spread of the tick along climate-determined trajectories and the emergence of Lyme disease in Canada were documented (Berry et al., 2014a; Smith et al., 2014; Beard et al., 2016; Ebi et al., 2018) (Table 6.1). These assessments also identified risks of the introduction of exotic vector-borne diseases, such as malaria, dengue, and chikungunya, with climate change. Increasing temperatures were anticipated to increase the geographical extent of North America suitable for transmission of exotic vector-borne pathogens and the survival of populations of the exotic mosquito vectors (e.g., Aedes species) that transmit them.
Table 6.1
Summary of assessment findings related to the effects of climate change on infectious diseases, excluding water- and food-borne diseases
Assessment
Identified climate-sensitive infectious diseases
Anticipated impact of climate change
Evidence for impacts of climate change on disease risks
Adaptation and adaptive capacity at the time of the assessment
Chapter 5: The Impacts of Climate Change on Water-, Food-, Vector-, and Rodent-Borne Diseases in Human Health in a Changing Climate: A Canadian Assessment of Vulnerabilities and Adaptive Capacity (Charron et al., 2008)
Tick-borne disease
Mosquito-borne disease
Hantavirus
Plague
Northward geographic range spread of tick-borne and mosquito-borne diseases into Canada from the United States
Invasion of exotic mosquito-borne diseases (dengue, malaria)
Invasion of exotic directly transmitted infections (e.g., SARS)
None
Adaptive capacity in terms of risk assessment, surveillance, prevention, and control is robust in Canada, but gaps identified in knowledge of disease ecology, the effects of climate, expert capacity, surveillance, and warning systems
Chapter 7: Human Health in Canada in a Changing Climate: Sector Perspectives on Impacts and Adaptation (Berry et al., 2014a)
As above
As above
Evidence for the emergence of Lyme disease via climate-determined trajectories, as well as increasing incidence of human cases
As above
Chapter 11: Human Health: Impacts, Adaptation, and Co-Benefits in Intergovernmental Panel on Climate Change (IPCC) Assessment Report 5 (AR5) (Smith et al., 2014)
Mosquito-borne diseases, such as malaria and dengue, in low- and middle-income countries
Tick-borne diseases in Europe and North America
Plague in Asia and North America
Hantavirus in North America
Possible increased incidence and range expansion of mosquito-borne and tick-borne diseases
None identified
Need to address risk and adaptive capacity in all countries by:
Reducing poverty
Improving nutrition, basic public health, and health services
Vulnerability mapping
Developing early warning systems linked to control programs
Chapter 26: North America in IPCC AR5 North America Chapter (Romero-Lankao et al., 2014)
Mosquito-borne diseases such as West Nile virus
Tick-borne diseases, particularly Lyme disease
Possible increased incidence and range expansion of mosquito-borne and tick-borne diseases
Risk of invasion of exotic mosquito-borne diseases
Evidence for the emergence of Lyme disease via climate-determined trajectories in Canada
Need improved datasets and models to understand effects of environmental changes versus other determinants of vector-borne disease risk, and early warning systems
Chapter 5: Vector-Borne Diseases in The Impacts of Climate Change on Human Health in the United States: A Scientific Assessment (Beard et al., 2016)
Mosquito-borne disease such as West Nile virus and dengue
Tick-borne diseases such as Lyme disease
Plague
Geographic range change, change in seasonality, and date of onset/duration of vector activity
None identified
Need improved models to understand and predict the effects of weather and climate changes versus other determinants of vector-borne disease risk, and field observations to support these
Chapter 14: Human Health in Impacts, Risks, and Adaptation in the United States: Fourth National Climate Assessment. (Ebi et al., 2018)
Mosquito-borne disease such as West Nile virus and dengue
Tick-borne diseases such as Lyme disease
Changes in geographic ranges, seasonal distribution, and abundance of disease vectors
Increased risks from mosquito-borne diseases abroad, which may spread into US territory
None identified
Need vulnerability and adaptation assessments and response plans, integrated surveillance for vector-borne diseases, and early warning systems such as disease forecasting
6.1.3
Managing Infectious Disease Risks
Share
Adapting to climate-related infectious diseases first involves assessing risks to health and identifying populations at increased risk to the impacts. The capacity of public health systems to detect emerging and re-emerging infectious diseases by surveillance, and to prevent and control them by health promotion or more proactive measures such as vaccine development, as well as the capacity of health care systems to minimize consequences of infectious diseases, need to be assessed. Assessments provide information on vulnerability to infectious diseases and identify the most effective measures to reduce it (Berry, 2008). In general, previous assessments have suggested that, in North America, the ability of public health systems to assess risks from emerging and re-emerging infectious diseases, and to detect, prevent, and control them, is robust (see Table 6.1). The risk to most Canadians is low because of the relatively high socio-economic status of much of the population, which provides an environment that limits many disease risks, and because of the strong public health and health care infrastructures and systems. In addition, progress has been made in filling gaps in knowledge, surveillance, capacity, and early warning systems identified in previous assessments (see section 6.4 Adaptation to Reduce Health Risks).
6.2
Methods and Approach
Share
This chapter includes analysis of the impacts of climate change on human risks from infectious diseases of importance for public health in Canada, with the exception of infectious diseases transmitted in drinking and recreational water and in food, which are addressed in separate chapters (see Chapter 7: Water Quality, Quantity, and Security, and Chapter 8: Food Safety and Security). This chapter is a narrative review authored by subject matter experts. However, to support the author team, a rapid review was conducted to identify the majority of the national and international literature on weather, climate, and climate change impacts on infectious diseases. The review focused on the five areas explored in the chapter:
vector-borne diseases:
exotic mosquito-borne diseases, including those for which humans are the main reservoirs (e.g., malaria, dengue)
mosquito-borne diseases endemic in Canada (e.g., WNV disease)
infectious diseases directly transmitted human-to-human (e.g., influenza and enterovirus infections)
infectious diseases transmitted by inhalation from environmental sources (e.g., cryptococcosis, legionellosis)
Canadian capacity to adapt to changing risks from infectious diseases
The rapid review followed the general framework for scoping reviews first proposed by Arksey and O’Malley (2005) and further refined over the last 15 years (Levac et al., 2010; Peters et al., 2015; Tricco et al., 2016). It identified and characterized all of the available international research on climate change and infectious diseases using a systematic and reproducible methodology. A protocol was developed a priori and defines the scope of the rapid review, the comprehensive search strategy, and all tools used to screen citations and extract information from the literature (available upon request). The electronic search was conducted using Embase, PubMed, and Global Health in September 2018 to identify relevant literature in English and French on infectious diseases that also explored effects of weather, climate, and/or climate change. A grey literature search included targeted hand-searching of various government and scientific websites to identify reports that were not indexed in the electronic databases. Search results were de-duplicated in the reference management software Endnote (EndNote X7, Clarivate Analytics), and unique citations were uploaded into the web-based systematic review software Distiller SR (Distiller SR, Evidence Partners, Ottawa, Canada). Studies were screened for relevance by two reviewers, working independently, using a relevance screening tool developed a priori. All citations that were included after relevance screening were procured and confirmed to be relevant, and then the research was characterized using the developed data characterization tool. More recent publications were identified during the chapter review process.
Where sufficient information exists, in this chapter the confidence in direction and strength of the effects of climate change are identified. Identification of climate- or weather-sensitivity of diseases was considered to provide limited evidence for effects of climate change, evidence for climate- or weather-sensitivity combined with projections of effects of climate change provides medium evidence, while detected changes in infectious disease risks attributable to recent climate change was considered as providing robust evidence for effects of climate change.
6.3
Climate-Sensitive Health Risks, Projected Impacts of Climate Change, and Evidence of Impacts to Date
Share
6.3.1
Vector-Borne Diseases
Share
6.3.1.1 Effects of Climate Change on the Ecology and Epidemiology of Vectors and Vector-Borne Disease Transmission
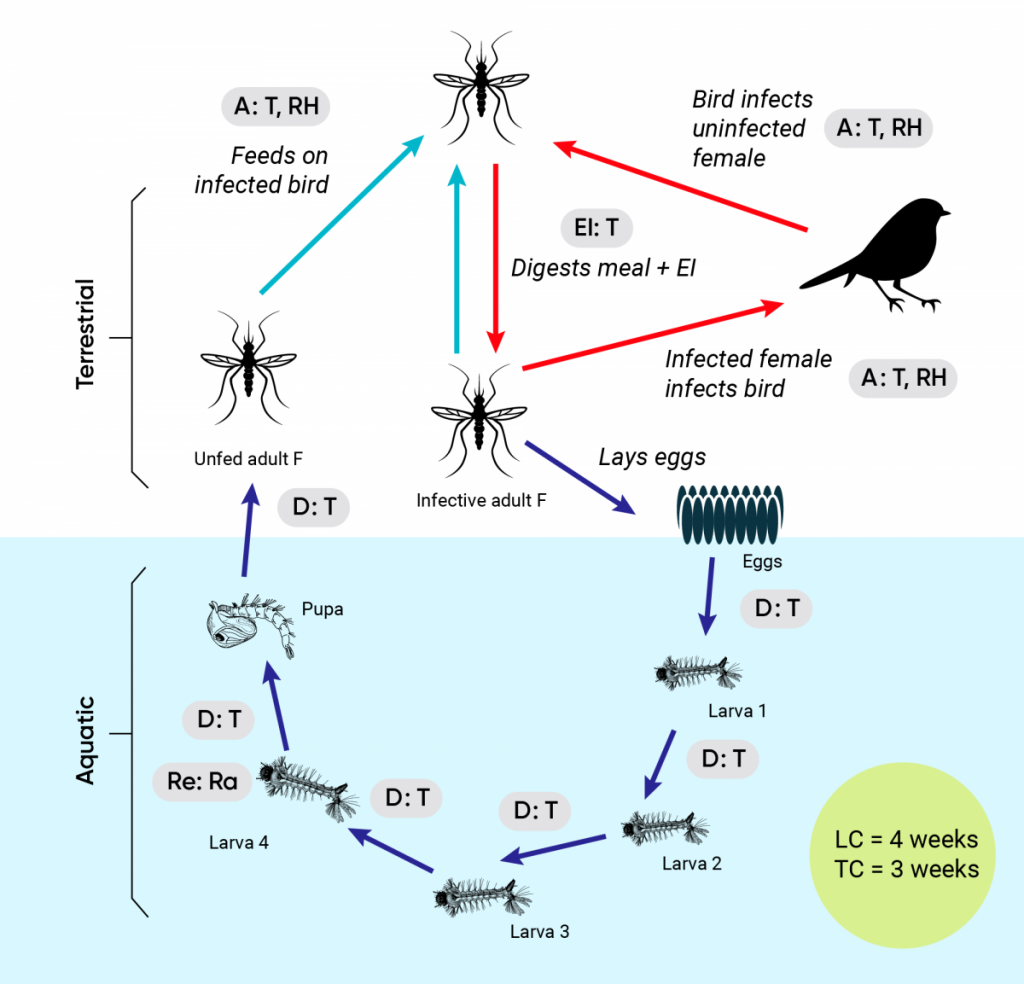
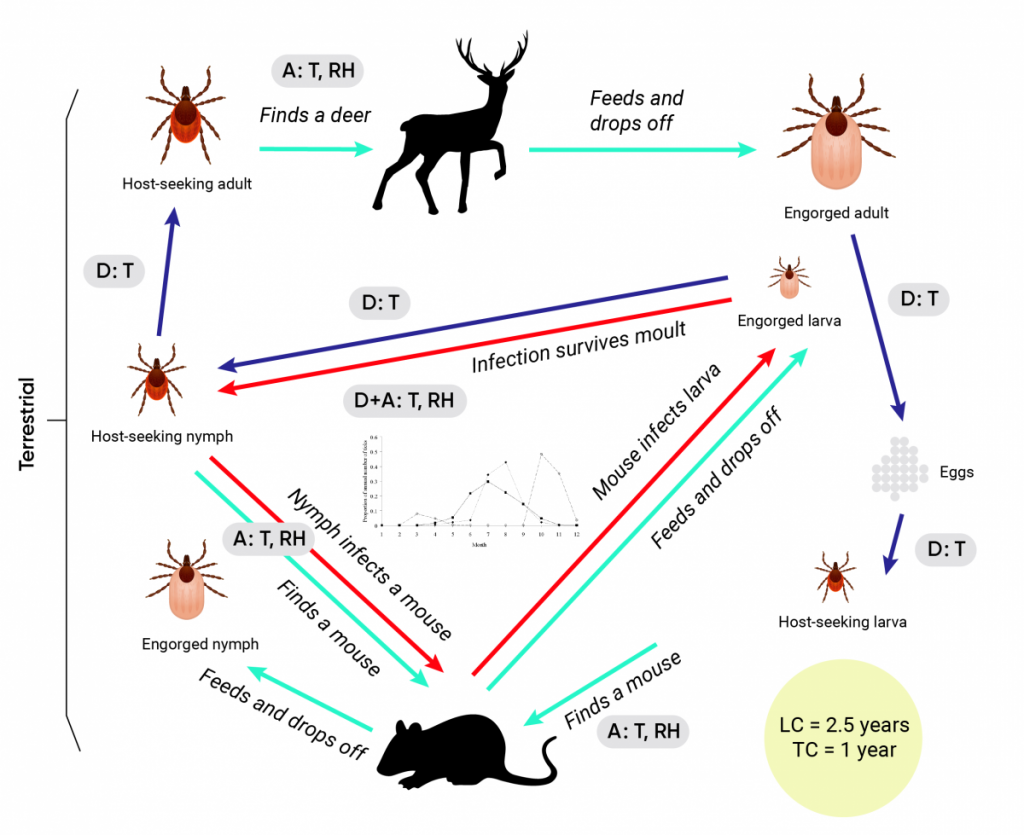
The life cycles of many arthropod vectors, and the impacts of weather and climate on these life cycles, have been intensely studied in the laboratory, field, and modelling studies, and reviewed elsewhere (Ogden & Lindsay, 2016). How changes in weather and climate may affect arthropod vectors and the transmission of vector-borne diseases is summarized in Box 6.1. While the effects of weather and climate are generic for arthropod vectors, how specifically they affect life cycles and transmission cycles is highly idiosyncratic among the different vectors and pathogens (Figure 6.3). Climate change may affect the risk of vector-borne diseases by altering human social-behavioural risk factors, such as perception of risk and adoption of preventive behaviours (Bouchard et al., 2018). Human population growth, movement, and social and economic factors (e.g., changing exposure to tick-borne encephalitis virus in eastern Europe due to changing population exposure following collapse of the Eastern Bloc) have also been associated with differential rates of human exposure to vectors and vector-borne diseases (Randolph, 2004). How climate change may affect risks indirectly through these factors requires further study.
Figure 6.3
Share
The impacts of weather and climate on the life cycles of dipteran and ixodid tick vectors.
1 / 2
Figure 6.3
The impacts of climate change are illustrated using the life cycle of the mosquito Culex pipiens and its role in the transmission of West Nile virus (WNV) in A. The life cycle of the tick Ixodes scapularis and its role in the transmission of the Lyme disease agent Borrelia burgdorferi is illustrated in B. In both, dark and pale blue arrows indicate development, and host finding/detachment from hosts, respectively, while red arrows indicate pathogen transmission cycles. Points at which weather and climate (and potentially climate change) may affect the vector’s life cycle (LC) and pathogen transmission cycle (TC) are indicated by the grey-filled boxes in which A = effects on activity, D = effects on inter-stadial development rates, EI = effects on the extrinsic incubation period, Re = effects on reproduction, T = effects of temperature, Ra = effects of rainfall, and RH = effects of humidity. In the centre of the B. burgdorferi transmission cycle (B) is an illustration of the seasonal activity of the tick vectors, which may be affected by climate and climate change through effects on both development and activity, as described in the text.
Source
Ogden & Lindsay, 2016.
6.3.1.2 Exotic Mosquito-Borne Diseases
There is a wide range of mosquito-borne diseases of public health significance globally. Some, such as malaria, dengue, yellow fever, and chikungunya, which are transmitted from human to human by mosquitoes, are among the world’s greatest public health threats, causing millions of cases and deaths annually (WHO, 2019a). Other diseases, many of which are mosquito-borne zoonoses, such as those caused by Sindbis and Venezuelan equine encephalitis viruses, occur more sporadically (Table 6.2). Exotic mosquito-borne diseases are absent from Canada due to multiple factors, including climate. For most exotic mosquito-borne diseases, Canada’s climate is currently too cold for the vectors, and/or for development of the pathogens in the mosquito (Ng et al., 2019). Other barriers exist, including the lack of animal reservoir hosts for some diseases (such as primates for yellow fever virus) and standards of housing, including doors and windows, which prevent entry of vectors, and air conditioning systems, which inhibit human-to-human transmission by mosquitoes (Reiter, 2001). Physical barriers need to be surmounted for the pathogens and vectors to be transported to Canada from overseas.
The expansion of the geographic range of exotic vector-borne diseases into Canada requires movement of exotic vectors and pathogens from the countries where they are endemic. Increasing global trade and travel are expected to facilitate the global movement of mosquito vectors (infected or not) and infected travellers, thereby increasing the possibility of importation of vectors and pathogens into regions where they have not previously occurred (Tatem et al., 2006; Tatem et al., 2012; Semenza et al., 2016). There are three scenarios whereby vector-borne diseases currently exotic to Canada may emerge here (Box 6.2).
The public health impact of climate change on each emergence scenario will vary and could include increased likelihood of Canadians acquiring infection while travelling abroad, increased likelihood of short-lived autochthonous (i.e., local, within Canada) transmission where competent vectors are already established and permanent endemicity of new diseases in Canada (Ng et al., 2019).
Mosquito vectors of exotic mosquito-borne diseases are sensitive to weather conditions, and climate change would be expected to affect them. Field and laboratory experiments demonstrate the temperature sensitivity of the Aedes and Anopheles species mosquitoes that are the main vectors of the most important exotic mosquito-borne diseases globally (Brady et al., 2013; Shapiro et al., 2017). In general, warmer temperatures, high humidity, and increased precipitation facilitate the life cycle of mosquitoes by supporting larval development and survival and by extending adult lifespan (Reeves et al., 1994; Jetten & Focks, 1997; Paaijmans et al., 2009; Yang et al., 2009). These climatic conditions will influence pathogen transmission by:
reducing egg development time in adult female mosquitoes, thus reducing time between blood feeds and increasing feeding frequency (Reeves et al., 1994; Jetten & Focks, 1997; Paaijmans et al., 2013);
shortening the EIP, thereby allowing mosquitoes to become infectious faster (Davis, 1932; Reeves et al., 1994; Jetten & Focks, 1997; Paaijmans et al., 2009; Paaijmans et al., 2013; Xiao et al., 2014); and
increasing mosquito longevity, enabling infectious mosquitoes to bite more people (Yang et al., 2009).
However, many of these weather-dependent relationships are complex; relationships can be non-linear, can have opposite effects depending on circumstance, and can be influenced by non-climatic factors (Box 6.1).
The degree to which climate change will affect, or has already affected, the global distribution of many mosquitoes and mosquito-borne diseases is uncertain. This is because the relationship between climate and vector-borne diseases, particularly those transmitted human-to-human via mosquitoes, is complex, and the spatiotemporal distribution of vectors and the pathogens they carry depends on a range of factors that are not directly climatic. These factors include increased mobility and interconnectivity of people and goods (Junxiong & Yee-Sin, 2015; Semenza et al., 2016; Tabachnick, 2016; Lindsey et al., 2018; Romeo-Aznar et al., 2018), urbanization and other land-use changes (Junxiong & Yee-Sin, 2015; Jones & O’Neill, 2016; Semenza et al., 2016; Asad & Carpenter, 2018; Romeo-Aznar et al., 2018), socio-economic factors (KC & Lutz, 2017; Reina Ortiz et al., 2017; Lindsey et al., 2018; Moreno-Madriñán & Turell, 2018; Romeo-Aznar et al., 2018), demographic changes, including those due to immigration and population growth (Asad & Carpenter, 2018; Lindsey et al., 2018; Romeo-Aznar et al., 2018), population immunity (Anyamba et al., 2012; Larrieu et al., 2014; Semenza et al., 2016), genetic evolution and adaptation (Gubler, 2007; Tsetsarkin et al., 2007; Tabachnick, 2016), access to health care (Reiter, 2008; Ooi & Gubler, 2009; Semenza et al., 2016), and vector control and intervention programs (Githeko et al., 2000; Junxiong & Yee-Sin, 2015; Tasanee et al., 2015). Some of these factors are independent of climate change, but some may be influenced by climate change, and climate change may affect mosquito-borne diseases indirectly via these factors.
Despite the uncertainty, effects of climate change on the spread of mosquitoes and mosquito-borne diseases are likely (high confidence), with regional variation across the globe (Smith et al., 2014). By shortening the length of the life cycle, increasing mosquito survival, and enhancing pathogen transmission, climate change will drive the expansion of mosquito and mosquito-borne pathogen populations in some locations and increase the geographic scope, mostly poleward and toward higher altitudes, of their ecological niches in many cases (Campbell et al., 2015; Kraemer et al., 2015; Samy et al., 2016; Hertig, 2019; Kamal et al., 2019; Kraemer et al., 2019). While mosquito populations may expand into new geographic areas, they may disappear from others (Machado-Machado, 2012; Escobar et al., 2016; Williams et al., 2016).
A warming climate is expected to enhance populations of Canada-endemic mosquito species (Hongoh et al., 2012) and conditions for pathogen transmission, making Southern Canada, in particular, more suitable for the emergence of new mosquito-borne diseases by scenarios 1 and 2 (Box 6.2). Temperature conditions in parts of Southern Canada are also expected to become increasingly suitable for populations of the yellow fever mosquito Ae. aegypti and the Asian tiger mosquito Ae. albopictus that are vectors of dengue, chikungunya, Zika, and yellow fever. Climate change is also expected to increase temperature suitability for virus transmission (Ng et al., 2017), rendering mosquito-borne disease emergence by scenario 3 (Box 6.2) increasingly possible in Southern Canada. Southern Coastal British Columbia, Southern Ontario, Quebec, New Brunswick, and Nova Scotia are anticipated to become climatically suitable for Ae. albopictus (Ogden et al., 2014a), while Southern Coastal British Columbia may also become suitable for Ae. aegypti (Campbell et al., 2015; Kamal et al., 2019). In 2016, an adult Ae. aegypti was collected in Southern Ontario. A very small number of adults and larvae were found in the subsequent year under enhanced mosquito trapping and field surveillance in the area, suggesting this species has not become established in Canada (Windsor-Essex County Health Unit, 2019a). However, Ae. albopictus has been found sporadically in multiple, but restricted, locations in Southern Ontario since 2005. There is one small area of Canada where Ae. albopictus became established between 2017 and 2018, although, to date, there is no evidence that these mosquitoes carry pathogens that have caused illness in humans (Windsor-Essex County Health Unit, 2019b).
Particularly for scenarios 2 and 3, the likelihood that new vector-borne diseases emerge in Canada will depend on the number of pathogen and/or vector introduction events, as well as the degree to which climate change makes Canada a more suitable environment for them to become established. Some forms of global movement may be driven by climate change, for example, climate refugees (McMichael et al., 2012) and changes in travel patterns (World Tourism Organization & United Nations Environment Programme, 2008). Climate change may therefore have three impacts that increase the likelihood of emergence by scenarios 2 and 3:
increased climatic suitability in Canada;
increased abundance of exotic pathogens and their vectors in countries outside Canada; and
increased introduction of pathogens and vectors.
Even without the effects of climate change, global interconnectivity is increasing (Findlater & Bogoch, 2018), and Canadians are enthusiastic travellers (Statistics Canada, 2019), so local and global movement will continue to pose a growing risk for the introduction of exotic pathogens via imported vectors and infected humans into Canada. Pathways of introduction may be predictable, however. For Japanese encephalitis virus (JEV), the most probable scenario for introduction into the United States would be by JEV-infected female mosquitoes arriving on aircraft from eastern China (Oliveira et al., 2018). JEV incursion would likely follow a similar pathway into Canada, with coastal British Columbia being the most likely entry location because of its many travel and trade connections with Asia. It also has a temperate climate and endemic mosquito vectors and reservoirs that might support local JEV transmission, should it arrive.
In addition to a suitable climate, other factors such as poverty and a lack of access to infrastructure, health care, and disease control measures are also important for endemic transmission of exotic pathogens transmitted human-to-human by mosquitoes (Ebi et al., 2006a; Halstead, 2008). Social conditions for most of the Canadian population (particularly population density and housing) are expected to limit mosquito biting rates, and health services remove infected people from transmission, making it difficult for efficient, sustained transmission cycles of such exotic pathogens to become established (Berrang-Ford et al., 2009; Ng et al., 2019). It is more likely that the public health impact will be occasional autochthonous cases in Canadians who have not travelled. These people may have severe health consequences because, in the absence of a history of travel, diagnosis may be delayed (Berrang-Ford et al., 2009; Ng et al., 2019). Delayed diagnosis of “unexpected” cases of exotic mosquito-borne disease in Canadians could lead to some chains of transmission and limited outbreaks. Examples from outside Canada include the introduction of chikungunya virus into Mediterranean Europe in 2007 and dengue in France, Croatia, and Madeira between 2010 and 2013.These events may have been driven by particularly warm weather conditions (Rezza et al., 2007; Tomasello & Schlagenhauf, 2013).
In contrast, if exotic mosquito-borne zoonoses invade, they may be more likely to become endemic if environmental conditions, including climate, are suitable, as pathogen transmission among wildlife and/or livestock reservoir hosts would be mainly unrestricted, as has been the case for WNV. The range expansion of Sindbis virus (SINV) and Usutu virus (USUV), out of Africa and into Europe, have strongly paralleled that of WNV in North America (Weissenböck et al., 2002; Ashraf et al., 2015). Given that the mosquito vectors and reservoirs for USUV and SINV are already present in Canada, and the climate in invaded parts of Europe is very similar to that of southern regions of Canada, the emergence of SINV or USUV in this country as disease-causing endemic viruses is possible. Infections with SINV and USUV produce unpleasant but relatively mild infections in immunocompetent humans.
Another exotic mosquito-borne zoonosis, Rift Valley fever virus (RVFV), can cause infections that are serious and can be fatal (WHO, 2019b), so the possibility of emergence of this virus in Canada is a more concerning threat. Canada-endemic mosquito species have been shown to be competent vectors for RVFV, while a wide range of wildlife and domesticated livestock are animal reservoirs (Box 6.2, Table 6.2). The most likely mechanism of introduction into North America is considered to be RVFV-infected humans arriving on aircraft from endemic areas where there is an outbreak (Golnar et al., 2014; Golnar et al., 2018). Should the traveller come into contact with endemic mosquito vectors, transmission to wildlife and/or livestock may result in endemicity.
Table 6.2
Key global mosquito-borne diseases and features of their transmission (vectors and hosts), geographic occurrence, and the presence of vectors and hosts in Canada and the United States
Disease
Pathogen(s)
Geographic distribution of human cases
Endemicity in the US
Primary vectors
Vectors established in Canada?
If absent from Canada, are vectors established in the United States?
Not endemic but locally acquired cases reported in 2014–2015 in Florida and Texas
Aedes aegypti and Ae. albopictus
Emerging population of Ae. albopictus in a very limited area of Southern Ontario
Aedes aegypti southern states to Southern New York state, Ae. albopictus in southern and northeast states, including those bordering Central and Eastern Canada
Humans, wild primates (in Africa)
Yes (humans), no wild primates
Yes
2
Dengue
Dengue virus serotypes 1 to 4
Asia, the Pacific, Americas, Africa, Caribbean
Endemic in Puerto Rico
Ae. aegypti and Ae. albopictus
Emerging population of Ae. albopictus in a very limited area of Southern Ontario
Aedes aegypti southern states to Southern New York state, Ae. albopictus in southern and northeast states, including those bordering Central and Eastern Canada
Humans, wild primates in Southeast Asia and Western Africa
Yes (humans), no wild primates
Yes
2
Japanese encephalitis
Japanese encephalitis virus
Asia, Pacific Islands, Torres Strait of Australia, Papua New Guinea
Not endemic
Culex spp. mosquitoes
Cx. tarsalis in Western and Central Canada
Domestic pigs and wild birds
Yes
Yes
2
La Crosse encephalitis
La Crosse virus
United States (upper mid-western and mid-Atlantic and southeast states)
Endemic to most of the eastern states, including states that border Canada
Aedes triseriatus
Aedes triseriatus in Eastern Canada
Small mammals (chipmunks and squirrels
Yes
Yes
1
Malaria
Plasmodium spp.
Central America, South America, Africa, Southeast Asia, Middle East, Southwest Pacific
Not endemic
Anopheles spp. mosquitoes, particularly An. gambiae and An. arabiensis
Anopheles quadrimaculatus in Eastern Ontario and Quebec, Anopheles freeborni in British Columbia
Humans
Yes
Yes
2
Rift Valley fever
Rift Valley fever virus
Eastern and Southern Africa, Saudi Arabia, Indian Ocean
Not endemic
African floodwater Aedes spp. mosquitoes
Some Aedes species in Canada are capable of transmitting the virus in the laboratory
Unknown, possibly livestock, equines, and wild ungulates
Yes
Yes
3
Sindbis fever
Sindbis virus
Northern Europe, Australia, China, South Africa
Not endemic
Ornithophilic Culex, Culiseta, Ochlerotatus. and Aedes spp. mosquitoes
Culex pipiens and some Ochlerotatus and Aedes spp.
Birds
Yes
Yes
2
St. Louis encephalitis
St. Louis encephalitis virus
Central, Western, and Southern United States
Endemic in northern, eastern, and central states including states bordering Canada
Culex tarsalis, C. pipiens, C. quinquefasciatus, C. nigripalpus
Culex tarsalis in Western and Central Canada and Culex pipiens along the US-Canada border
Birds
Yes
Yes
1
Usutu virus
Usutu virus
Africa, Europe
Not endemic
Culex pipiens, Cx. neavei
Culex pipiens along the US-Canada border
Birds
Yes
Yes
2
Venezuelan equine encephalitis
Venezuelan encephalitis virus
South and Central Americas, Mexico and cases as far as Southern United States
Not endemic
Culex, Aedes, Mansonia, Psorophora, Deincerites, and Ochlerotatus spp. mosquitoes
Oc. sollicitans in Eastern Canada
Oc. taeniorhynchus along most of US coast and Culex melanoconion in Florida
Rodents, equines
Yes
Yes
2
Yellow fever
Yellow fever virus
Tropical and subtropical Africa, South America
Not endemic
Ae. aegypti
Aedes aegypti southern states to Southern New York state
Humans, wild primates
Yes (humans), no wild primates
Yes
3
Zika
Zika virus
Africa, Southeast Asia, South America
Not endemic but locally acquired cases in Puerto Rico, Florida, and Texas from 2015–2016
Ae. aegypti, Ae. albopictus
Emerging population of Ae. albopictus in a very limited area of Southern Ontario
Aedes aegypti southern states to southern New York state, Ae. albopictus in southern and northeastern states, including those bordering Central and Eastern Canada
Humans, wild primates
Yes (humans), no wild primates
Yes
2
6.3.1.3 Canada-Endemic Mosquito-Borne Diseases
All four of the most medically important arboviruses endemic to Canada — WNV, EEEV, snowshoe hare virus (SSHV), and Jamestown Canyon virus (JCV) — are transmitted through bites of infected female mosquitoes. Mosquitoes acquire infections from specific mammalian or avian reservoir hosts. The main mosquito vectors for WNV are Cx. pipiens and Cx. restuans in Eastern Canada and Cx. tarsalis in Western Canada (Kramer et al., 2008), while for EEEV, Culiseta melanura is the main vector (Armstrong & Andreadis, 2010). Non-Culex mosquito species (e.g., Aedes, Culiseta, and Anopheles spp.) are the primary vectors of the California serogroup viruses (CSGV), such as SSHV and JCV (Drebot, 2015; Pastula et al., 2015; Webster et al., 2017). For WNV and EEEV, a wide range of bird species serve as reservoirs, including corvids and passerines (Kilpatrick et al., 2006; Kramer et al., 2008; Ludwig et al., 2010; Reisen, 2013). The main reservoir of JCV is the white-tailed deer (Andreadis et al., 2008), while squirrels, chipmunks, and hares are the reservoir hosts for SSHV (Drebot, 2015). A number of these viruses are also maintained by transovarial transmission, which allows for less dependence on mammalian reservoirs (Griot et al., 1993).
Additional viral and bacterial agents transmitted by insects are also endemic in Canada but are less active, or their occurrence is under-studied. Western equine encephalitis virus appears to have decreased in prevalence in Canada in recent decades, while Cache Valley virus (CVV) has been responsible for a number of livestock (i.e., sheep) outbreaks in Ontario, Quebec, and other provinces, but human infection is most likely under-reported (Drebot, 2015). Arboviruses can also be transmitted occasionally by blood transfusion or tissue transplants (Fonseca et al., 2005; Pathogen Regulation Directorate, 2010). Apart from this possibility, humans are incidental/dead-end hosts for these mosquito-transmitted diseases; while they can be infected, they cannot subsequently transmit viruses to feeding mosquitoes with any efficiency because viremia is transient and viral loads are low (Kramer et al., 2008; Kulkarni et al., 2015).
Approximately 20% of individuals who are exposed to mosquito-borne viruses, such as WNV, EEEV, JCV, or SSHV, will develop acute clinical illness, including fever, headache, skin rash, nausea, and muscle aches. Most affected people recover fully, but approximately 1% develop severe illness (e.g., meningitis, encephalitis, acute flaccid paralysis, and poliomyelitis), in which case neurologic and cognitive deficits may be prolonged or permanent. Approximately 10% of severe cases are fatal. Individuals over 70 years of age and those with underlying medical conditions, such as obesity, diabetes, hypertension, and heart disease, are at greater risk of severe illness. However, SSHV causes neurological illness in children as well. People who are immunocompromised are also at greater risk (Petersen et al., 2013a; Petersen et al., 2013b; Sejvar, 2014; Badawi et al., 2018). The severity of illness varies and depends upon the virus; for example, EEEV is one of the most severe mosquito-transmitted diseases in the United States, with approximately 33% mortality in those developing neurological illness and significant brain damage in most survivors who developed symptomatic disease (Centers for Disease Control and Prevention, 2018c). Western equine encephalitis virus and CVV give rise to a similar range of symptoms; while the majority of cases are asymptomatic, a varying percentage develop encephalitis, meningoencephalitis, encephalomyelitis, high fever, altered consciousness, neurologic dysfunction, aseptic meningitis, stiff neck, headache, myalgia, tremors, nausea, vomiting, and urinary tract infection. The mortality rate is between 5% to 20% for St. Louis encephalitis virus, but is believed to be much lower for western equine encephalitis virus and CVV infection (Centers for Disease Control and Prevention, 2018d).
The expected effects of climate change on Canada-endemic mosquito-borne diseases are northward range expansion associated with long-term warming and more epidemic behaviour associated with climate variability and extreme weather events, via effects on vector survival and reproductive rates (together affecting vector abundance), biting rates, the length of the activity season, and the duration of the EIP. Canada-endemic mosquito-borne diseases are zoonoses transmitted from wild animals (birds and mammals). The effects of climate change on the populations of these animals are expected to affect the pathogen transmission cycles. These effects may simply result in northward range expansion of hosts, but there may be more complex effects on reservoir host biodiversity. For example, changes in host abundance and geographic range may be limited by physical conditions (e.g., barriers to movement) and/or biological processes (e.g., reduced access to food at critical times in the life cycle, such as breeding and rearing periods). Resulting changes in species composition can have varying consequences, such as disruptions in predator–prey and host–parasite relationships. Therefore, although host biodiversity will likely change in response to new climate conditions, uncertainties remain regarding how such changes will affect exposure risk of Canadians to vector-borne diseases (Varrin et al., 2007). In all likelihood, the impact will be specific to the ecosystem or habitat, resulting in a patchwork of increasing and decreasing biodiversity of host communities, changing with time across the country. Climate change may have more rapid effects on host communities via extreme weather events, such as droughts and heat events, which can bring reservoir hosts searching for water sources to mosquito breeding grounds (Shaman et al., 2005; Wang et al., 2010; Harrigan et al., 2014).
Many modelling studies have examined the relationship between climatic variables (mainly temperature and precipitation) and WNV infection (infected humans, birds, or mosquitoes) in Canada (Wang et al., 2011; Chen et al., 2013; Tam et al., 2014; Paz, 2015; Yoo et al., 2016; DeFelice et al., 2018). However, the ecology of EEEV and CSGV remains under-studied, likely due to perceptions that these viruses are not as important for public health, as well as a lack of detailed surveillance data. However, land cover, including proximity and size of coniferous forested area and wetlands, has been found to influence EEEV and JCV occurrence (Vander Kelen et al., 2014; Rocheleau et al., 2018) and could also be affected by climate change.
The impact of climate change on WNV transmission in Canada has been investigated in two studies with similar conclusions. Chen et al. (2013) examined WNV transmission in the Prairies, where Cx. tarsalis is the main vector, and projected an extension of seasonal activity of WNV-infected Cx. tarsalis from three months (June to August) to five months (May to September) by the 2080s. The authors also projected a northward range expansion for Cx. tarsalis and WNV. Since this vector is also capable of transmitting CVV, the range and prevalence of this disease may also be influenced by this Cx. tarsalis expansion (Ayers et al., 2018). Hongoh et al. (2012) modelled the potential distribution of Cx. pipiens populations in Eastern Canada under current and future projected climate change and projected a similar northward range expansion for this eastern vector of WNV.
A greater understanding of how climate change may alter communities of avian and mammalian reservoir host species would enable more robust evaluation of climate change effects, but few studies have conducted these evaluations, due to limited data and methodological constraints. Current evidence indicates that climatically suitable ranges (or climate envelopes) for many species will likely shift northwards in response to warming temperatures. For example, ecological niche models for 765 species suggest that climate change may increase biodiversity in Southern Quebec during this century, as species move northward (Berteaux et al., 2010; Chambers et al., 2013). Similarly, many bird species that currently breed in the northern portion of the Eastern United States are likely to move northward into Canada, increasing the richness of bird species in Eastern Canada (DesGranges & Morneau, 2010). Habitat loss and disturbance, induced by climate change or other factors, that may result in habitat fragmentation (Warren & Lemmen, 2014) can affect avian and mammalian reservoir host communities (Berteaux & Stenseth, 2006). To what extent these positive and negative effects on host populations will cause increased or decreased risks from mosquito-borne diseases is not yet clear and needs further study (Salkeld et al., 2013).
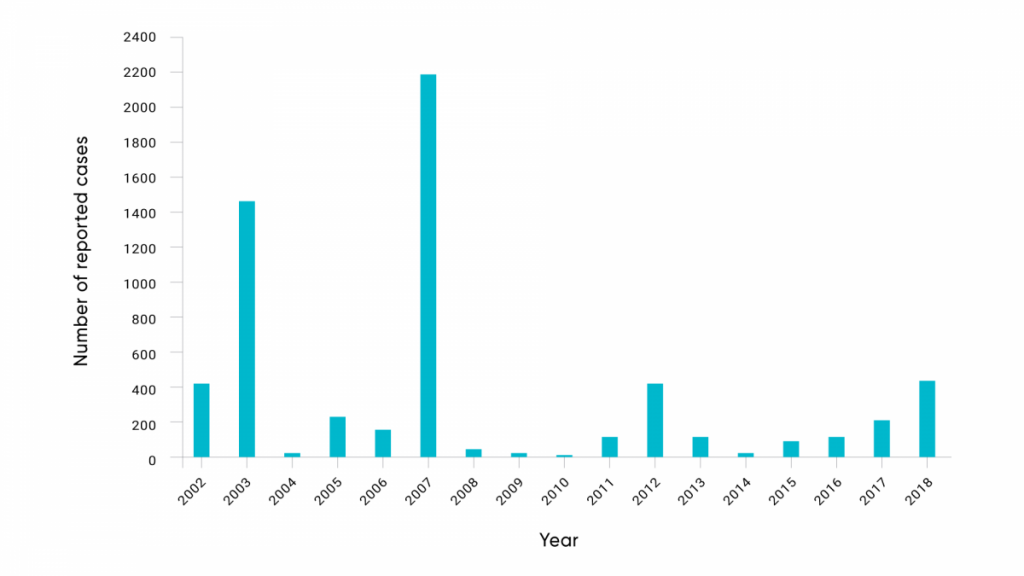
Mosquito-borne infections have been identified in Canada for many decades; however, recently the number of cases of arbovirus infection appears to be increasing (Ludwig et al., 2019). Since 2002, the annual reported incidence of human cases of WNV, the only Canada-endemic mosquito-borne disease that is nationally reportable, has fluctuated significantly over time at a national level. Reported cases have ranged from five cases in 2010 to highs of 1481 in 2003 (during initial invasion across Canada) and 2215 cases in 2007, associated with an unprecedented abundance of Cx. tarsalis mosquitoes in the Prairie provinces (Figure 6.4). This may be consistent with the effects of weather and climate variability on WNV dynamics (Ludwig et al., 2019).
The number of reported human cases of WNV each year in Canada.
Source
Government of Canada, 2019a.
Geographical variation over time has been dramatic as well. In 2003 and 2007, most human cases of WNV were reported in the Prairies (Alberta, Saskatchewan, and Manitoba), but in 2002, 2012, and 2018 most reported cases were detected in Ontario and Quebec. This variability is at least in part consistent with the effects of local weather variability on the abundance of Cx. tarsalis,Cx. pipiens, and Cx. restuans mosquitoes during outbreak years, and an indication that greater weather variability under climate change may result in more epidemic behaviour of endemic mosquito-borne diseases (Ludwig et al., 2019). Human CSGV cases have been detected across Canada, and a single human EEEV case was reported in 2016 in Ontario (M. Drebot, personal communication, 2019).
Increased awareness of CSGVs, enhanced field surveillance in reservoir hosts, and greater diagnostic capacity in humans and animals, may have contributed to their “emergence” as a public health concern during the mosquito season. Routine diagnostic testing for CSGVs was conducted during the late 1970s and 1980s but discontinued until new testing methods were introduced in 2005, when human cases were once again documented in Canada. Over 200 probable and confirmed cases of CSGV infections and/or exposures have been identified by laboratory-based surveillance from 2005 to 2014, with illness from JCV being more frequently detected than illness caused by SSHV (Drebot, 2015; Lau et al., 2017; Webster et al., 2017; M. Drebot, personal communication, 2019). Although CSGV infections are not nationally notifiable, the numbers of CSGV infections have been summarized in the Public Health Agency of Canada’s (PHAC) arbovirus annual reports (Government of Canada, 2019a) and have ranged from 34 to 122 cases per year. To date, there have been no direct associations observed between the effects of weather variability or recent climate change and the incidence of these mosquito-borne viruses in Canada, although such associations may exist.
In Canada, changes have been observed in the geographic distributions and densities of mosquito vectors. The mosquito fauna of Canada, which includes 74 mosquito species from 10 different genera, was described in the 1970s (Wood et al., 1979). Since then, six species (Ochlerotatus ventrovittis, Oc. japonicus, Culex salinarius, Culex erraticus, Anopheles perplexens, and An.crucians) have been reported as possibly newly established in Canada (Thielman & Hunter, 2007; Giordano et al., 2015; Iranpour et al., 2017). In addition, the geographic range of 10 species (Uranotaenia sapphirina, Culiseta melanura, Cs. minnesotae, Culex tarsalis, Ochlerotatus sticticus, Oc. spencerii, Oc. dorsalis, Oc. nigromaculis, Oc. campestris, and Oc. cataphylla) has expanded in Canada (Iranpour et al., 2009). Some of these range expansions, which may affect public health, could have been facilitated by changes in climate, but lack of systematic surveillance precludes any conclusion.
There is strong observational evidence of range shifts for mammal and bird species in North America. Over the past 40 years, about 180 of 305 bird species wintering in North America expanded their range northward, at an average rate of 1.4 km per year. Similarly, the breeding ranges of birds in Southern North America have shifted by an average of 2.4 km per year (Federal, Provincial, & Territorial Governments of Canada, 2010). Within the northeastern forests of North America, 27 of 38 species for which historical ranges are documented have expanded their ranges, predominantly northward (Rodenhouse et al., 2009). Published accounts of range shifts in Canada are available for a number of species (Hitch & Leberg, 2007; Blancher et al., 2008;), with detailed analyses for some species, including the hooded warbler (Setophaga citrina) (Melles et al., 2011), and the southern flying squirrel (Glaucomys volans) (Garroway et al., 2010; Garroway et al., 2011). It is very possible that geographic range shifts of these species have been driven, in part, by recent climate warming. Such range changes could affect endemic mosquito-borne pathogen transmission by changing ranges of reservoir species, while species that are not reservoirs may act to “dilute” arbovirus transmission cycles (Levine et al., 2017). However, further study is required to understand precisely how, where, and when this may have an impact on risk to humans.
6.3.1.4 Other Insect-Borne Zoonotic Diseases
Other insect-borne zoonotic diseases may also be affected by climate change. Plague, caused by the bacterium Yersinia pestis, has been documented sporadically in Western Canada. Yersinia pestis is transmitted by the bite of an infected flea or by direct contact with infectious tissues or fluids while handling an animal or human that is sick with, or has died from, plague. Droplet-transmission via coughing or sneezing is also possible, due to infection of the lungs of an animal or human with pneumonic plague (Centers for Disease Control and Prevention, 2019a). People infected with plague usually develop flu-like symptoms. After this flu-like phase, they develop varying symptoms, depending on the form of plague — bubonic, septicemic (this form typically develops as a complication of bubonic plague), or pneumonic. Plague is an infection that requires urgent medical care, as mortality rates are high in the absence of treatment (Centers for Disease Control and Prevention, 2019a). The natural reservoirs of Y. pestis are wild rodents, particularly ground squirrels in North America and gerbils in Asia.
There is a well recognized association of climatic patterns (generally warmer and wetter periods) with spillover of plague from gerbil–flea transmission cycles to humans in central Asia, due to effects on vegetation that promote gerbil and then flea populations (Kausrud et al., 2010; Samia et al., 2011). These spillover events in Asia are thought to have been the source of the great plague pandemics (the Justinian plague and the Black Death) that decimated human populations in Europe (Kausrud et al., 2010). The impact of climate was studied on plague outbreaks in pre-industrial Europe (1347 to 1760 CE). In contrast to spillover in the Asian steppes, the results suggested that plague in Europe was associated with drier and colder climates (Yue & Lee, 2018). This difference is likely due to the transmission in Europe being driven by a combination of direct human-to-human transmission (causing pneumonic type of plague in humans) and flea-borne transmission from peri-domestic rat reservoirs (producing the bubonic type of plague), both of which may be influenced by effects of climate on human population density and behaviour (Earn et al., 2020). A 56-year time series of human plague cases in the Western United States was used to explore the effects of climatic patterns on plague incidence. As in central Asia, warmer and wetter climate was associated with increased numbers of human cases (Ben Ari et al., 2008). In a consecutive study, the same group found that El Niño–Southern Oscillation and Pacific Decadal Oscillation, in combination, affect the dynamics of plague over the Western United States by enhancing dry-to-wet changes in the climate. The underlying mechanism could involve changes in precipitation and temperatures that affect both hosts and vectors as in Asia. Snow may play a key role, possibly via effects on summer soil moisture, which affects flea survival and development and growth of vegetation for rodents (Ben Ari et al., 2010). A study of the relationships between climatic variables and the frequency of human plague cases (from 1960 to 1997) in Northeastern Arizona and Northwestern New Mexico suggested that plague risk can be estimated by monitoring key climatic variables, most notably maximum daily summer temperature values and time-lagged (one- and two-year) amounts of late winter (February–March) precipitation (Enscore et al., 2002).
Modelling studies have been conducted on the impact of climate change on plague distribution in North America. Using an ecological niche-modelling approach, models by Holt et al. (2009) suggest that, by 2050, climate conditions may reduce plague risk in the southern parts of California and increase risk along the northern coast and the Sierras. A study by Nakazawa et al. (2007) suggested that the disease shifts in accordance with patterns of climatic shift, but that overall geographic shifts will likely be subtle, with some northward movement of southern limits and possibly northward movement of northern limits as well.
Studies of flea species vectors of Y. pestis in Canadian prairie dog populations suggested that flea counts per individual varied inversely with the number of days in the prior growing season with more than 10 mm of precipitation; an index of the number of precipitation events that might have caused a substantial, prolonged increase in soil moisture and vegetative production (Eads & Hoogland, 2017). Beyond these studies there have been no attempts to assess how precisely climate change may impact plague dynamics and geographic range in Canada, and there are no field surveillance data available to explore any climatic impacts on the environmental hazard of plague in Canada. Plague is nationally notifiable, but only one human case has been recorded, which occurred in 1939 (Government of Canada, 2018a).
Chagas disease is caused by the protozoal parasite Trypanosoma cruzi and is an infection most commonly acquired through contact with the feces of an infected triatomine bug (or “kissing bug”), a blood-sucking insect that feeds on humans and animals. Chagas disease has an acute and a chronic phase and, if untreated, infection is lifelong. Infection may be mild or asymptomatic. There may be fever and/or swelling around the site of inoculation. Many people may remain asymptomatic for life, but 20% to 30% of infected people develop debilitating and sometimes life-threatening medical problems over the course of their lives (Centers for Disease Control and Prevention, 2018e). It is absent from Canada and, while most of the estimated 300,000 cases of Chagas disease in persons living in the United States were likely acquired in Latin American countries, transmission cycles of T. cruzi involving animal hosts and humans, and autochthonous vector-borne human infections, have been reported in Texas, California, Tennessee, Louisiana, and Mississippi in the United States (Steverding, 2014).
Chagas disease is likely under-recognized and having an impact on the health care system and economy because of limited screening and treatment and a lack of awareness among health care professionals (Click Lambert et al., 2008; Bern & Montgomery, 2009). The possible impact of climate change on Chagas disease vectors in North America has been explored using ecological niche-modelling methods; while northward range expansions of some species were predicted, increasing risks for Canada were not found (Carmona-Castro et al., 2018).
6.3.1.5 Tick-Borne Zoonotic Diseases
Ticks transmit a wide range of bacterial, viral, and protozoan pathogens globally (Sonenshine, 2018). While it is generally acknowledged that increases in temperature associated with climate change will likely contribute to a general increase in the number, type, activity level, and geographical distribution of ticks in North America (Eisen et al., 2016; Sonenshine, 2018), the magnitude of impact climate change will have on risks from tick-borne diseases is uncertain and will likely vary regionally. In Canada, evidence suggests that the emergence of Lyme disease, associated with the northward range spread of the tick Ixodes scapularis, has been driven, at least in part, by recent climate warming (Ebi et al., 2017; Hoegh-Guldberg et al., 2019).
Tick-borne diseases of public health significance are zoonoses, and in North America the natural reservoir hosts are wild animals, particularly rodents. There are two types of ticks, hard-bodied (Ixodid) ticks and soft-bodied (Argasid) ticks (Lindquist et al., 2016). In Northern North America, including Canada, the soft-bodied tick of most public health importance is Ornithodorus hermsi, which transmits the bacterium that causes relapsing fever, Borrelia hermsii. Other soft-bodied ticks and relapsing fever Borrelia species occur in the United States; Borrelia turicatae and Borrelia parkeri are transmitted by Ornithodoros turicata and Ornithodoros parkeri, respectively, but are not currently endemic to Canada (Sage et al., 2017). Both O. hermis and B. hermsii are naturally cave-dwelling species, and the natural hosts for blood meals by the ticks, and reservoir hosts for B. hermsii, are wild rodents. While Ornithodorus hermsi and B. hermsii most commonly occur in caves in the mountainous regions of the Western United States and have limited distribution in Southern British Columbia, they invade cabins in these regions, which is where most human infections are acquired (Dworkin et al., 2008).
Tick-borne relapsing fever is a febrile, septicemic disease with a sudden onset followed by numerous relapses with afebrile intervals (Artsob, 2000; Murray, 2003). Persistence of the bacterium and relapses are associated with bacteria evading the immune response (Cutler, 2010). There is a very wide range of symptoms, including rashes, ocular lesions, jaundice, and vomiting (Ogden et al., 2014b). However, it is uncommon in humans in Canada, due to the low frequency with which people come into contact with infected ticks.
An ecological niche-modelling approach identified elevation (higher elevations being more favourable) and specific ranges of temperature and precipitation as key determinants of the presence of O. hermsi ticks and B. hermsii (Sage et al., 2017). In this same study, wider northward and westward range expansion into mountainous regions of British Columbia were projected using three global climate models (GCM) (GCMs: ACCESS1-0, HadGEM2-ES, and CCSM4) and two estimates of greenhouse gas concentration trajectories denoted by representative concentration pathways (RCP) 4.5 and 8.5. Greater range expansion was projected in Canada with the higher greenhouse gas emission scenario, RCP8.5. For both RCP4.5 and RCP8.5, all models projected a range contraction in the United States.
Diseases transmitted by hard-bodied ticks pose the greatest tick-borne disease challenges for public health in North America, and, among these, the most important is Lyme disease, caused by the bacterium Borrelia burgdorferi. Lyme disease is a disease affecting multiple body systems that begins with mild non-specific illness and, in most cases, a typical skin rash known as erythema migrans. If untreated, the disease progresses to disseminated Lyme disease with neurological or cardiac manifestations and, in late stages, arthritis (Wormser et al., 2006). This bacterium is transmitted by Ixodes scapularis (the black-legged tick) in Northeastern and Upper Midwest United States, and in Southern Central and Eastern Canada, and I. pacificus (the western black-legged tick) in the Pacific states of the United States and Southern British Columbia (Bouchard et al., 2015; Eisen et al., 2016). In Southern British Columbia, the geographic range of I. pacificus, and the risk of B. burgdorferi infection the tick poses, is thought to be quite wide (Mak et al., 2010). However, the risk of acquiring Lyme disease is much lower where I. pacificus is the vector, compared to most regions where I. scapularis is the vector, due to characteristics of the ecology of I. pacificus that result in generally low infection prevalence in this tick and less likelihood that it bites humans (Eisen et al., 2016). There have been no studies to date to assess possible impacts of climate change on future I. pacificus and B. burgdorferi distributions in British Columbia. Modelling of current I. pacificus and B. burgdorferi distributions in British Columbia has identified temperature — specifically, mean daily temperatures in January and July — as an important determinant of the ecological niche of these species, and climate change is expected to have an impact on northward and possibly altitudinal distributions (Mak et al., 2010).
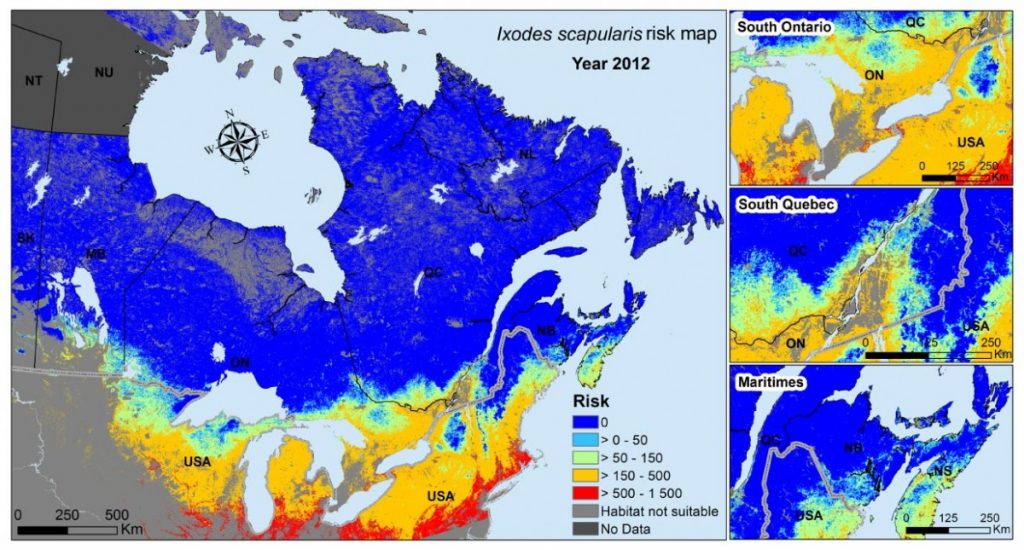
For I. scapularis, a range of field and laboratory studies (Ogden, 2014) suggested that the main impact of temperature on this species is on development rates and activity; woodland habitats in Canada provide refuges for the ticks, in which they are protected from the direct effects of very low winter temperatures that would otherwise kill them. Model-based assessments of the risk of occurrence of I. scapularis used effects of temperature on development rates, and thus life cycle length, to identify lower temperature limits for persistence of self-sustaining tick populations (Ogden et al., 2005). These limits have now been extensively validated by field studies, which, in concert with analysis of passive tick surveillance data, have identified a spatiotemporal pattern of south-to-north range spread into Canada from the United States (and now within Canada) that is consistent with recent climate warming having been a key driver (Ogden et al., 2010; Leighton et al., 2012; Clow et al., 2017; Ebi et al., 2017).
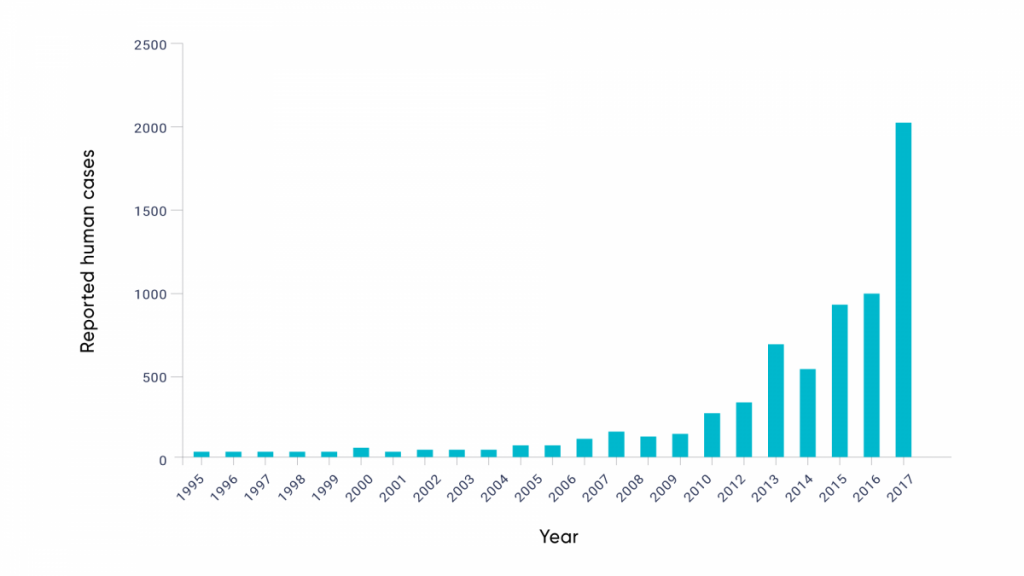
The rapidly increasing incidence of Lyme disease in Canada, identified by national surveillance (Gasmi et al., 2017), is consistent with the observed range expansion of I. scapularis, as well as increasing infection prevalence in recently established tick populations (Ogden et al., 2013; Clow et al., 2017) (see Figure 6.5). Due to the high level of agreement among studies, and the evidence of climate change impacts, there is high confidence that the emergence of Lyme disease in Eastern and Central Canada has been associated with recent climate warming. The observed emergence of Lyme disease in Canada is consistent with effects of climate change acting on the tick vector itself. However, there is also evidence that a warming climate may be influencing the risk of Lyme disease via effects on other parts of the transmission cycle, particularly the abundance and geographic range of key rodent reservoir hosts (Simon et al., 2014).
The evolution of Lyme disease risk in Canada and human cases.
Figure 6.5
Note: Bar chart showcases the evolution of human Lyme disease cases. Note that Lyme disease became nationally notifiable in December 2009. Data from 2009 and earlier are based on voluntary submission of information from provinces and territories in which Lyme disease was notifiable.
Source
Gasmi et al., 2018.
Other Ixodes scapularis-transmitted pathogens, additional to B. burgdorferi, are now emerging in Canada from the United States as the ticks spread north (Table 6.3), but none of the diseases they cause are currently nationally notifiable. Details of these are presented in Box 6.3. A model-based assessment of the possible effects of climate change on the geographic range of the tick Dermacentor variabilis, a vector of RMSF and tularemia, which is already endemic to much of Southern Canada, suggested that the range of this tick will expand northward in Canada (Minigan et al., 2018).
Prolonged extreme values of temperature (i.e., high or low), low humidity, and intense rainfall could adversely affect tick survival by reducing their activity and increasing their mortality rate (Ogden & Lindsay, 2016). However, this is expected to have less of an effect on ticks than on mosquitoes, because of the tick’s ability to find refuge in their woodland habitats (Ogden & Lindsay, 2016). Model-based studies have suggested that, while the northern limit of the geographic range of I. scapularis and D. variabilis ticks will move northward with climate change, there will also be a northward contraction of the southern limit of the ticks’ range as the climate becomes too hot for them (Brownstein et al., 2005a; Minigan et al., 2018). If so, this may affect exposure to the ticks for those living in southern US states, but is unlikely to affect risk in Canada. Studies of I. scapularis suggest that ticks in the Southeastern United States may adapt to a hot climate (Ogden et al., 2018), so this effect may be minimal.
Non-climate factors also determine where tick populations and pathogen transmission cycles can become established. These include microhabitat features, such as soil characteristics, which are critical for tick survival and the successful establishment of new tick populations (Lindsay et al., 1998; Guerra et al., 2002). Modifications in habitat characteristics, in parallel with climate change, such as habitat fragmentation, loss of biodiversity, resource availability, and land use, affect the dynamics of ticks, their animal hosts, and the exposure of ticks to humans (Brownstein et al., 2005b; Simon et al., 2014). In some parts of Canada, the emergence of I. scapularis and B. burgdorferi is patchy and uneven, likely because of the suitability of different woodlands for tick survival and because of variations in tick host abundance (Gabriele-Rivet et al., 2015). Changes in non-climate factors can drive emergence of tick-borne diseases. For example, Lyme disease emerged in the United States in the 20th century, likely as a consequence of the reforestation of farmland and the increase in the deer populations, which allowed the expansion of the tick populations (Kilpatrick et al., 2017).
While Lyme disease and other tick-borne diseases transmitted by I. scapularis and I. pacificus ticks are the tick-borne diseases of greatest concern for public health in Canada at present, other Canada-endemic tick-borne diseases (Table 6.3) are possibly being affected by a warming climate or may be affected in the future. Currently, there is no evidence of the impact of recent warming on these diseases; however, there is evidence from surveillance showing recent increases in the abundance and range of tick vectors of these other diseases. These include increased abundance of I. cookei (a vector of Lineage I Powassan virus), Dermacentor variabilis (a vector of RMSF, tularaemia, and Colorado tick fever), and Rhipicephalus sanguineus (a vector of RMSF) in Eastern Canada (Gasmi et al., 2018), and range expansion of D. variabilis in the Prairie provinces (Dergousoff et al., 2013) (Table 6.3).
The northward range expansion of Lyme disease into Canada provides proof of concept of the following:
The geographic range of ticks and tick-borne diseases in North America may be limited by climate.
A warming climate may permit northward range expansion of these species.
Migratory birds and, perhaps, other mammalian tick host species act as a route of introduction of ticks and tick-borne pathogens each year from the United States into parts of Canada that are becoming climatically suitable for their endemicity (Ogden et al., 2008; Nelder et al., 2019).
Therefore, other US-endemic ticks and tick-borne pathogens that are not currently endemic to Canada may spread north. Northward expansion of the range of Amblyomma americanum, a vector of tularemia, Heartland virus, and the bacterial pathogens Ehrlichia ewingii and E. chaffeensis, as well as the suspected cause of red meat allergies (Reynolds & Elston, 2017), has been detected in the United States (Stafford et al., 2018). Multiple model-based assessments suggest that areas of Southern Canada will become climatically suitable for this tick with a warming climate (Raghavan et al., 2019; Sagurova et al., 2019). This tick, and the infectious disease and other public health issues it brings, are likely to spread into Canada in the coming decades, likely introduced by migratory birds (Gasmi et al., 2018; Nelder et al., 2019).
Table 6.3
Tick-borne pathogens affecting humans and their associated vector tick species that occur in Canada or may spread into Canada
Eastern, central, western, and southwestern states
No
Yesb
Yes
Yes
Source: Bouchard et al., 2019
Note: * Where there is no specific disease name, diseases are named after the pathogen (e.g., Powassan virus disease)
(–) indicates no data available and/or no studies have been performed
a Canada: Provinces where endemic transmission is known to occur are underlined. For provinces that are not underlined, pathogen transmission cycles are unknown, and infections were detected in adventitious ticks, and/or in humans or animals most likely infected by adventitious ticks.
US: States where highest incidence rate of human cases were found.
b Based on historical surveys in ticks in Canada, rather than recent surveys.
c Francisella tularensis may be mechanically transmitted by a range of biting flies, but only ticks act as biological vectors.
Abbreviations: AB, Alberta; BC, British Columbia; MB, Manitoba; NB, New Brunswick; NL, Newfoundland and Labrador; NS, Nova Scotia; ON, Ontario; PEI, Prince Edward Island; QC, Quebec; SK, Saskatchewan; US, United States; NE, Northeast; MW, Midwest.
6.3.2
Zoonoses Directly Transmitted from Animals to Humans
Share
In this section, zoonoses that can be transmitted directly from animals to humans are considered (Figure 6.6). Zoonoses transmitted in food and water are considered in Chapters 7: Water Quality, Quantity, and Security, and Chapter 8: Food Safety and Security. The risk of infection by directly transmitted zoonoses may be influenced by direct effects of climate change and weather on the survival of the pathogens, indirect effects on their host species and communities, and/or effects on contact rates between the pathogens and humans. Together, these determine the frequency of “spillover” events, that is, when pathogens are successfully transmitted from animals to humans (Altizer et al., 2013; Brierley et al., 2016), as well as the possibility of adaptive emergence of zoonoses, which then result in epidemics or pandemics affecting human populations (Ogden et al., 2017). Climate change may, therefore, drive global emergence of zoonotic infectious diseases. Directly transmitted zoonoses, mostly from wildlife, are the zoonoses that emerge and re-emerge most frequently on a global scale (Jones et al., 2008).
Climate change impacts on zoonoses in the North are expected to be greater than elsewhere in Canada, including effects on vector-, food-, and water-borne zoonoses, as well as directly transmitted zoonoses (Parkinson et al., 2014). Greater climatic impact, coupled with, in many cases, higher consumption of traditional and country foods,1 suggest that Northern communities, particularly Northern Indigenous communities, are at greater risk of health effects (Brook et al., 2009). A further concern is that, as permafrost melts with a warming climate, pathogens that have remained dormant but viable in animal carcasses or soil, may be released into the environment and cause disease outbreaks (Revich et al., 2012). For most of the directly transmitted zoonoses discussed in this section, there is evidence of sensitivity to climate and/or weather, which raises the possibility of current and future impacts of climate change. In very few cases have there been attempts to assess such impacts. Also, for very few directly transmitted zoonoses is there evidence of changing patterns of disease or risk, but that may be due, in part, to the lack of systematic surveillance of these diseases in Canada.
Zoonoses and emergence by genetic change and/or evolution.
6.3.2.1 Viral Zoonoses
Globally, one of the most highly pathogenic and high-impact groups of directly transmitted viral zoonoses are avian influenza A viruses. Global epidemics of highly pathogenic (for humans) avian influenza viruses have occurred in recent decades (Goneau et al., 2018), and outbreaks have occurred in domesticated poultry in Canada, sparking fears of human cases (Skowronski et al., 2007). Disease in humans varies from mild illness to severe illness, with high case-fatality rates (Neumann, 2015). The main reservoirs of avian influenza viruses are wild birds, particularly waterfowl (i.e., swans, geese, and ducks). Influenza viruses of swine are also of great public health concern, although these viruses are now maintained mostly by domesticated swine, and effects of climate change are relatively unlikely (Schultz-Cherry et al., 2011). Pandemic influenza viruses, such as the 1918 and 2009 H1N1 viruses, emerge following recombination (a molecular process) of avian, swine, and human influenza viruses (Neumann et al., 2009). Due to the high level of threat from zoonotic and pandemic influenza viruses, there is extensive surveillance for them in wild and domesticated animals and humans, both globally and in Canada (Government of Canada, 2017; Government of Canada, 2018b), and a program of preparedness for pandemics (Government of Canada, 2019b).
Emergence and re-emergence of avian influenza risks to humans could be driven by a number of climate change effects on the ecology of the viruses, including bird migration patterns and changes in land use and livestock production patterns (Morin et al., 2018). Transmission of avian influenza viruses among wild birds, which often involves fecal–oral transmission via water, may be inhibited by climate warming, as the survival of the viruses is greater in cool compared to warm water (Morin et al., 2018). Consistent with this, the prevalence of infected birds in a Canadian study was higher if sampling was preceded by cold weather (Papp et al., 2017), and phylogenetic data suggest that influenza A viruses actually emerged following the Little Ice Age (Gatherer, 2010). However, complexity associated with the global transmission of avian influenza viruses means that, while avian influenza viruses are inherently climate-sensitive, the effects of climate change could be positive, with reduced disease transmission because of effects on environmental survival of the viruses, or negative, with increasing transmission through indirect effects of other factors (Gilbert et al., 2008; Gatherer, 2010; Morin et al., 2018).
Rabies is a zoonotic disease that represents a major worldwide health concern with more than 55,000 human deaths annually (Knobel et al., 2005). Rabies virus is capable of infecting cerebral and nervous tissues of all mammals, usually leading to behavioural changes followed by death (WHO, 2013). In Canada, rabies is associated with several wildlife species, including bats, skunks, raccoons, and foxes, that each maintain distinct viral variants circulating endemically within their populations (Rosatte, 1988; Tinline & Gregory, 2020). Human cases of rabies are rare in Canada, with only three domestically acquired cases since 2000 (Filejski, 2016), but human exposures to rabid animals followed by administration of post-exposure prophylactic treatment occur annually across the country.
While climate-driven ecological change is likely to affect the circulation of rabies in each of its respective host populations to some extent, such impacts are likely to be felt first and most significantly in the Arctic. Rabies occurs across the Arctic and is regarded as enzootic, or prevalent, in Northern Canada (north of 60 degrees north latitude) (see Chapter 2: Climate Change and Indigenous Peoples’ Health in Canada). The main reservoir host of Arctic rabies is the Arctic fox (Vulpes lagopus).
Arctic rabies is caused by a unique strain of rabies virus, referred to as the Arctic rabies virus variant, which circulates within the Arctic and sub-Arctic regions, with sporadic incursions toward more southern regions, occasionally leading to establishment of enzootic rabies in these areas (Rosatte, 1988). In the Canadian Arctic, rabies cases in domestic animals and wildlife are reported every year, and Arctic residents receive more rabies post-exposure prophylactic treatments per capita than any other population in Canada (Rosatte, 1988; Mitchell & Kandola, 2005; Aenishaenslin et al., 2014; CFIA, 2019).
Northern Canada is experiencing climate change at rates faster and greater than the global and Canadian averages (Larsen et al., 2014; Bush & Lemmen, 2019). While the long-term impacts of rapid climate warming on Arctic rabies have yet to be documented, rabies ecology and epidemiology are likely to be altered by ongoing climate-driven perturbations to Arctic ecosystems. These include fading population cycles of lemmings, a key source of food for Arctic foxes (Fuglei & Ims, 2008; Kausrud et al., 2008), reduced extent and duration of sea ice used by foxes for winter foraging and movement (Kim et al., 2016), and the northward range expansion of red foxes (Vulpes vulpes). Red foxes are both a resource competitor for Arctic foxes and competent reservoir host for the Arctic rabies variant (Rosatte, 1988; Hersteinsson & MacDonald, 1992; Gallant et al., 2012). A recent modelling study (Simon et al., 2019) suggested that rabies incidence may initially stabilize due to reduced variation in prey dynamics, with interactions between Arctic and red foxes intensifying outbreaks where these species overlap. However, in the long term, displacement of Arctic foxes by red foxes and restricted winter movements due to reduced sea ice extent may limit rabies transmission in warmer areas. Similarly, Huettmann et al. (2017) predicted a reduction of the current ecological niche of rabies virus in Alaska with climate warming.
Hantaviruses cause infections of wild rodent and insectivorous mammal populations and can cause hantavirus pulmonary syndrome, a fever followed by acute pulmonary edema and shock. There is no specific treatment for hantavirus pulmonary syndrome; the case-fatality rate is 38% in Canada (Drebot et al., 2000), where, on average, four cases are reported a year (Government of Canada, 2015). Humans become infected by contact with infected rodents or their excretions, particularly aerosolized urine or feces (Weir, 2005). In a previous report (Charron et al., 2008), the climate sensitivity of hantavirus incidence, associated with effects on rodent populations, was identified. There have been no further studies of hantavirus in Canada, and potential impacts of climate change are unexplored.
6.3.2.2 Parasitic Zoonoses
In Canada, the zoonotic protozoans Cryptosporidium, Giardia, and Toxoplasma are the most important parasites for public health, according to expert opinion, and the first two are nationally notifiable (FAO & WHO, 2014). Transmission of Cryptosporidium and Giardia spp. can be directly from animals, for example, from neonatal farm animals or in petting zoo environments. It can also occur from human to human through close contact and outbreaks in communal, caregiving environments, and through food or water. The effects of climate change on infections by these parasites will most likely occur through food- or water-borne transmission (see Chapter 7: Water Quality, Quantity, and Security, and Chapter 8: Food Safety and Security).
Currently, the human burden of disease, including neural and ocular lesions associated with visceral larval migrans (i.e., the migration of larvae through the body) due to the zoonotic ascarids (i.e., roundworms) Baylisascaris procyonis and Toxocara canis is not known in Canada, and only isolated cases and serosurveys are reported (Embil et al., 1988; Sapp et al., 2016). The reservoir hosts for these ascarids are, respectively, raccoons and domestic dogs. In Southern Canada, dogs are also competent hosts for B. procyonis (Lee et al., 2010). Toxocara canis was the most common parasite detected in the feces of dogs in shelters in a national study in Canada (Villeneuve et al., 2015). Direct transmission from dogs to humans is possible if eggs that have larvated in the environment stick to the fur of the animal, but in general T. canis is thought to be acquired mainly from eggs in the environment (Keegan & Holland, 2010).
Eggs of both ascarid parasites undergo temperature-dependent development in the environment before becoming infective for humans. Therefore, the primary mechanisms by which these parasites may be affected by climate change are through local amplification due to more rapid development and increased survival of eggs shed in the feces of definitive hosts (i.e., those in which parasite reproduction occurs), and through northward range expansion due to changes in the distribution and abundance of the canine and raccoon hosts. Eggs of T. canis are freeze-susceptible (O’Lorcain, 1995) and undergo delayed development after chilling (Azam et al., 2012), but eggs of B. procyonis can survive freezing temperatures and freeze–thaw cycles (Shafir et al., 2011). This suggests that a changing climate may have different effects on these parasites. For B. procyonis, northward expansion may follow that of the primary host, raccoons, which are currently limited to the southern parts of Canadian provinces (Naughton, 2012). For T. canis, northward range expansion and local amplification may follow increased survival and development rate of eggs in the environment. Increased circulation of both parasites in dogs, with their close relationship to people, may result in increased disease in Canadians. No studies to date have explicitly investigated effects of climate change on these parasites. However, eggs of T. canis were recently reported in dogs north of 60 degrees north latitude in Canada for the first time (Salb et al., 2008), which may suggest northward range expansion of this species (Jenkins et al., 2011).
Until recently, the zoonotic cestode Echinococcus multilocularis was thought to be transmitted between wild canids and rodents only in southern parts of the Prairie Provinces and the Arctic tundra, and only one autochthonous human case had been documented in Canada (James & Boyd, 1937; Deplazes et al., 2017). Eggs of Echinococcus are shed in the feces of wild or domestic canids; they are immediately infective and can adhere to the host’s fur. Infection can occur in humans via inadvertently consuming these eggs. Transmission can also occur via consumption of unwashed produce or drinking unfiltered surface water in heavily contaminated environments (Deplazes et al., 2017). In humans, the parasite establishes initially in the liver, and behaves like a parasitic tumour, eventually metastasizing throughout the abdomen unless aggressively surgically and medically managed. Recently, more cases of alveolar echinococcosis have been detected in both dogs and humans in Canada, and the parasite has expanded its range to include most of Western Canada (Deplazes et al., 2017) and the southern portion of Ontario (Kotwa et al., 2019). This emergence is likely linked to both the introduction of European strains of the parasite, which have now established successfully in wildlife, and increasing rates of contact between people and urban wildlife.
Possible mechanisms for the effects of climate change on this parasite include effects on egg survival, as eggs are environmentally resistant but susceptible to high temperatures and desiccation, and by stabilizing and amplifying rodent intermediate host populations, which were thought to be a limiting factor keeping this parasite in check (Rausch, 1956; Jenkins et al., 2013). Climatic variables, both temperature and precipitation, have been correlated with the prevalence of infection in definitive and intermediate hosts (i.e., hosts in which parasite reproduction does not occur) (Takeuchi-Storm et al., 2015), which may be associated with direct effects on the survival of eggs in the environment, or impacts on reservoir hosts (Mas-Coma et al., 2008). The parasite is now provincially notifiable in Alberta and Ontario. Dogs can serve as both definitive and intermediate hosts for E. multilocularis, and therefore regular cestocidal treatment of dogs known to consume rodent intermediate hosts may help mitigate human risk (Jenkins, 2017).
6.3.2.3 Bacterial Zoonoses
Brucella species bacteria, which are endemic to Canada and cause disease in humans, are Brucella abortus and B. suis biotype 4. Brucella abortus is maintained by cattle, when the disease is not prevented by control programs, and by wild ungulates, including deer and bison (Nishi et al., 2002). Brucella suis biotype 4 is maintained by caribou (Forbes, 1991). Brucellosis in humans causes persistent and often recurrent fever with chronic fatigue and other symptoms such as arthritis (Centers for Disease Control and Prevention, 2019b). In animals, the main effect of infection is abortion. The main transmission route to the human population is mostly food-borne, but direct transmission from tissues of infected livestock to those working in livestock and food-processing industries is possible (Hunter et al., 2015), as is direct transmission from infected tissues of wildlife to hunters (Franco-Paredes et al., 2017). Apart from possible direct effects of climate change on the survival of Brucella spp. bacteria in the environment (Aune et al., 2012), and thus on animal-to-animal transmission, climate change may affect transmission through effects on wild host population dynamics (Cross et al., 2007). While brucellosis may be a climate-sensitive disease, there have been no studies to explore possible effects of climate change.
Caused by the bacterium Bacillus anthracis, anthrax risk in the environment is amplified by infected wild and domesticated ungulates, which suffer hemorrhagic illness as a consequence of infection. The infected animal’s blood and carcass, if and when it dies, contaminates soils with bacteria that form resistant and persistent spores (Valseth et al., 2017). Humans can acquire infection by inhaling spores, ingesting spores contaminating food or water, or by contamination of skin cuts with spores.
Manifestations in humans depend on the route of infection — cutaneous manifestations after the infection of cuts, gastrointestinal manifestations after ingestion, and respiratory manifestations after inhalation. There is high case fatality (approximately 25%) in untreated people (Centers for Disease Control and Prevention, 2017). High temperatures have been associated with outbreaks in wild and domesticated animals in northern countries, including Canada, and enhanced environmental suitability for anthrax transmission and B. anthracis survival in the North and in South-Central Canada is anticipated with a warming climate (Walsh et al., 2018). The persistent nature of B. anthracis spores in soils allows weather-driven outbreaks to occur, and these may become more common with climate change due to combined effects of warming and flooding (Maksimovic et al., 2017). Both anthrax and brucellosis are reportable diseases when they occur in livestock in Canada (Government of Canada, 2019c).
Q fever is caused by the bacterium Coxiella burnetii. This bacterium is transmitted to humans mainly by airborne pathways from infected ruminant farms, particularly sheep and goat farms. In humans, the infection causes fever as well as pneumonia, infections of the liver, and chronic infections of the heart, and has a high case-fatality rate without treatment (up to 25%) (Centers for Disease Control and Prevention, 2019c). In ruminants, there are few clinical manifestations beyond abortion if the animal is infected in late pregnancy. Infected animals excrete the bacteria in milk, urine, feces, and placenta and birthing fluids (Plummer et al., 2018). The bacteria persist in the environment; while farm workers are those most at risk, large outbreaks have occurred in Europe associated with long-distance airborne transmission from affected farms (Schneeberger et al., 2014). Q fever is not nationally notifiable in Canada but is thought to be particularly prevalent in farms in Quebec (Dolcé et al., 2003), although the reason for this is unknown. Greater incidence has been associated with increased precipitation in the Caribbean, suggesting sensitivity to weather (Eldin et al., 2015; Sivabalan et al., 2017). The dispersal of the bacteria is increased with wind speed and low humidity, but reduced precipitation associated with climate change in some regions may reduce the concentration of this airborne pathogen (van Leuken et al., 2016). There are no studies exploring possible effects of climate change.
6.3.3
Infectious Diseases Directly Transmitted from Human to Human
Share
This section focuses on the subset of infections that are transmitted directly from human to human. Direct transmission can be defined as an infectious agent being transferred between people by direct contact or droplet spread rather than via food, water, or arthropod vectors. Direct contact occurs through skin-to-skin contact, whereas droplet spread refers to spray with relatively large, short-range aerosols produced by sneezing, coughing, or talking (Dicker et al., 2012). Directly transmitted infections cause a wide spectrum of illnesses, and many human population characteristics, such as changing demographics and human behaviour, patterns of connectivity, and compliance with public health recommendations (e.g., immunization against influenza), influence the burden of these diseases within the population (Heesterbeek et al., 2015).
The basic reproduction number (R0) (i.e., the number of secondary cases created by a primary disease case in a totally susceptible population and in the absence of intervention), is an important metric of communicability (Pandemic Influenza Outbreak Research Modelling Team, 2009). For directly transmitted pathogens, R0 is a function of duration of infectiousness, the effective contact rate of an infected individual, and the “infectivity” of an individual case (Pandemic Influenza Outbreak Research Modelling Team, 2009). If any of these factors are modified directly or indirectly by climate change, the result will be a change in disease epidemiology. Increases in R0 can increase the burden of endemic diseases and can make disease outbreak and emergence events more likely. In fact, the impacts of climate change on human habitation, agricultural productivity, conflict, livelihoods, and political stability will likely result in the displacement of populations and changes in mixing patterns that will have knock-on effects on communicable diseases; the link between human conflict, displacement of populations, and large-scale migrations and communicable diseases has been repeatedly demonstrated in recent and distant history (Rabaan, 2019). Such second- and third-order impacts of climate change on communicable diseases may dwarf the first-order changes in epidemiology due to impacts on hosts and pathogens.
There have been many studies identifying impacts of weather and climate on infectious diseases transmitted directly from human to human, but there is a dearth of studies that have attempted to explicitly investigate the effects of climate change. Only effects for hand-foot-and-mouth disease (HFMD) have been studied to date, and there have been no efforts to attribute any changes in disease incidence to recent climate change.
A possible indicator of climate-related modification of the epidemiology of directly transmitted communicable diseases is their seasonality. The seasonality of such infections is well recognized and has worked its way into the vernacular (e.g., “flu season”). While mechanisms underlying the seasonality of communicable diseases remain poorly understood (Fisman, 2007; Greer et al., 2008; Fisman, 2012), a growing body of literature has sought to distinguish the contribution of environmental factors to disease transmission, from other seasonally varying factors, such as seasonal human behaviour (school attendance, holidays, indoor crowding, etc.) and neuroendocrine factors (melatonin, vitamin D, etc.) (Dowell, 2001). Many viral and bacterial pathogens, particularly those that cause respiratory and gastrointestinal infection, display marked seasonality in temperate climates such as Canada’s, which suggests that environmental factors are important determinants of disease risk and environmental change may result in important changes in disease epidemiology.
6.3.3.1 Respiratory Infections
Respiratory infections are the greatest contributor to the overall burden of disease in the world (Ferkol & Schraufnagel, 2014) and were the third leading cause of death in 2015 worldwide (Global Burden of Disease, 2015; Mortality and Causes of Death Collaborators, 2016). Added to this now is COVID-19, but, to date, there is no evidence of the influence of weather and climate on the emergence and spread of this disease. Communicable respiratory diseases include diseases caused by both viral and bacterial pathogens. For many viral respiratory diseases, such as influenza, a broad spectrum of illness is common; most people suffer mild to moderate illness that is self-limited. However, a proportion of the population will experience more severe illness, or secondary bacterial infection, requiring hospitalization. It is estimated that, in Canada, an average of 12,200 hospitalizations (Schanzer et al., 2006; Schanzer et al., 2008; Schanzer et al., 2013a; Schanzer et al., 2013b) and approximately 3500 deaths are attributable to influenza annually (Schanzer et al., 2013b). Severe respiratory infections are most common at the extremes of age (Ampofo et al., 2006; Schanzer et al., 2006; Schanzer et al., 2008; Schanzer et al., 2013a; Schanzer et al., 2013b).
In temperate regions, viral respiratory infections, such as influenza and respiratory syncytial virus (RSV) can occur throughout the year, but seasonal epidemics peak in winter months (Thompson et al., 2006; Rohr et al., 2011). The predictable seasonality of wintertime viral respiratory infections has led investigators to evaluate whether environmental conditions could influence the time of onset and/or severity of seasonal influenza epidemics. There is some evidence that decreased absolute humidity (Shaman et al., 2010), decreased temperature (Earn et al., 2012; Skog et al., 2014), or both (Chattopadhyay et al., 2018) may trigger the onset of influenza season in temperate countries. Lower temperatures and decreased absolute humidity may also increase the transmissibility of influenza and influenza-like illness during epidemics (Tang et al., 2010; Roussel et al., 2016; Caini et al., 2018).
The severity of influenza seasons is diminished in the presence of strong El Niño–Southern Oscillation conditions (Viboud et al., 2004; Choi et al., 2006), providing further evidence of climate sensitivity. This thermal inversion in the Pacific Ocean is associated with unusually warm and rainy conditions, similar to those projected under climate change scenarios (Fisman et al., 2016). Given the apparent increase in influenza transmission in cold conditions, it might be expected that the effect of climate change would be to diminish the burden of illness associated with viral respiratory infections. However, climatic effects on influenza epidemiology may depend on time scale. An analysis of climate and past US influenza epidemic seasons from 1997 to 2013 found that mild winters were associated with early and severe epidemics in the subsequent year (Towers et al., 2013). Diminished influenza transmission in a given season may result in a larger population of susceptible individuals, and consequently a more explosive influenza epidemic, in the next season. Furthermore, it has been suggested that increased global temperatures may increase the rate of influenza diversification, which could diminish cross-protective immunity and give rise to more frequent influenza pandemics (Gatherer, 2010).
Several important bacterial respiratory human pathogens display stereotyped seasonality. Both Streptococcus pneumoniae and Neisseria meningitidis infections are more common in winter, which may be driven in part by increased risk due to prior influenza and/or respiratory syncytial virus infection (Tuite et al., 2010; Kuster et al., 2011). Direct environmental impacts on these infections are also possible. Invasive pneumococcal infections tend to be strongly seasonal in temperate climates, which leads to causal correlations with cold weather (Cilloniz et al., 2017). However, it has been suggested that diminished wintertime UV radiation, which is less likely to be influenced by climate change than temperature or precipitation patterns, may account for this wintertime seasonality (White et al., 2009). More recent modelling work suggests that pneumococcal seasonality may be complex and driven by interplays between population contact patterns, wintertime weather, and predisposing viral respiratory infections (Domenech de Cellès et al., 2019). Meningococcal infections have historically displayed highly seasonal occurrence in sub-Saharan Africa, with epidemic onset linked to the timing of the Harmattan winds (Sultan et al., 2005). In temperate regions, invasive meningococcal disease incidence often peaks in late winter or early spring (Rosenstein et al., 2001; Lindsay et al., 2002; Brachman & Abrutyn, 2009). However, links to environmental factors, while identified in individual regions, are inconsistent across regions (Collier, 1992; Lindsay et al., 2002; Sultan et al., 2005; Michele et al., 2006; Kinlin et al., 2009). Increased frequency of extreme weather events, including heavy rainfall events, are projected under climate change, and increases in respiratory illness have been associated with exposure to floodwaters (De Man et al., 2016). However, these illnesses are likely due to environmentally derived pathogens (e.g., Legionella, as described below) and water-borne pathogens, which are addressed elsewhere in this report (see Chapter 7: Water Quality, Quantity, and Security).
6.3.3.2 Gastrointestinal Infections
As with respiratory viral infections, gastrointestinal tract viral infections display highly seasonal occurrence. Enteroviral infections, including echovirus, coxsackie A and B viruses, and, before its control, poliovirus are associated with outbreaks in late summer and early autumn in temperate countries (Moore, 1982). Norovirus and rotavirus commonly occur in winter (Greer et al., 2009; Patel et al., 2013). Enteroviruses are usually transmitted via the fecal–oral route and can cause recognizable disease entities, including HFMD, neuroinvasive infections (e.g., acute flaccid paralysis), and non-specific syndromes, including viral meningitis, encephalitis, myocarditis, gastroenteritis, and conjunctivitis. Norovirus and rotavirus are associated with gastroenteritis, and norovirus in particular can cause large point-source outbreaks, including those in closed environments such as cruise ships and long-term care facilities (Rajagopalan & Yoshikawa, 2016; Mouchtouri et al., 2017).
Mechanisms underlying the seasonality of enteroviral infections are not fully understood. However, Dowell (2001) pointed out that the seasonality of polio becomes more evident with distance from the equator, and is absent peri-equatorially, implying that seasonal changes in weather are key drivers of enteroviral seasonality. It has been suggested that temperature modulates both viral reproduction and particle survival, and that humidity alters viral attachments to water droplets, facilitating transmission (Patz et al., 2005; Wong et al., 2010). Consistent with these hypotheses, elevated temperatures have been associated with increased HFMD risk in several Asian countries (e.g., Hii et al., 2011; Onozuka & Hashizume, 2011), at lags of one to two weeks. While some studies suggest that the relationship between temperature and disease risk is linear (Hii et al., 2011; Wang et al., 2016), or associated with high maximum daily temperatures (Sumi et al., 2017), others suggest more complex non-linear relationships between risk and temperature (Kim et al., 2016; Liao et al., 2016; Zhao et al., 2018).
Increased relative humidity and rainfall have also been associated with elevated HFMD risk in most studies (Hii et al., 2011; Onozuka & Hashizume, 2011; Zhao et al., 2017); humidity may modulate the effects of temperature (Kim et al., 2016; Sumi et al., 2017). It has been suggested that extreme precipitation events may enhance the risk of HFMD (Cheng et al., 2014; Yu et al., 2019). As noted above, the combination of increased temperature and heavy rains is associated with El Niño–Southern Oscillation, which are associated with near-term increases in HFMD in China (Lin et al., 2013). These observations have allowed parameterization of mathematical models that suggest a 5% increase in HFMD in China by 2090, which would represent a substantial increase in disease burden (Zhao et al., 2018). If such estimates are applicable to enteroviral infections generally, this could also present an increase in currently rare but severe manifestations of enteroviral disease, such as neuroinvasive infections and myocarditis.
Increased temperatures and rainy, humid weather associated with climate change could therefore increase the incidence of endemic enteroviral disease worldwide as well as the frequency of outbreaks, and spur the emergence of enteroviral strains in new geographic regions by elevating R0 beyond the threshold for sustained person-to-person transmission. Indeed, with the recent emergence of acute flaccid paralysis due to non-polio enteroviruses in North America, including Canada, such a phenomenon may currently be occurring (Hassel et al., 2015; Holm-Hansen et al., 2016; Suresh et al., 2018). Candidate vaccines against enterovirus EV-71, which is an important agent of HFMD and strongly associated with neuroinvasive disease, are in development but are not yet available for use in North America (Zhu et al., 2014).
In contrast to enteroviruses, both noroviruses and rotavirus strains are associated with wintertime gastroenteritis in North America (Greer et al., 2009; Patel et al., 2013). In Spain, unusually cold weather has been associated with increased risk of rotavirus-related hospitalization (Morral-Puigmal et al., 2018). While rotavirus is highly transmissible via person-to-person spread, environmental reservoirs (particularly water) may be important for spread, and lower water temperatures delay environmental decay of the virus (see Chapter 7: Water Quality, Quantity, and Security). Cold, dry conditions have also been associated with norovirus activity in England and Wales (Lopman et al., 2009). Consequently, climate warming in Canada could reduce the incidence of some of these viral pathogens.
6.3.4
Infectious Diseases Acquired by Inhalation from Environmental Sources
Share
Perhaps the best-known infectious diseases of humans that have environmental sources other than infected humans, animal reservoirs, or arthropod vectors, are legionellosis and cryptococcosis. These types of infections are caused by bacteria and fungi that persist in soils and water and that infect humans by inhalation after becoming aerosolized or airborne in dust. For all such infections, while there have been studies to identify influences of weather and climate on their occurrence, there has been little assessment of the possible impacts of climate change. Details of the pathogens, the diseases they cause, and their sensitivity to weather and climate are provided in the main text, while the possible effects of climate change are described in Box 6.5, Box 6.6, Box 6.7, Box 6.8, and Box 6.9.
6.3.4.1 Legionellosis
Infection with Legionella bacteria causes legionellosis, which presents as Legionnaires’ disease, Pontiac fever, or as an asymptomatic infection. Legionnaires’ disease manifests as pneumonia (Stout & Yu, 1997). Illness is often severe and can progress to respiratory and multi-organ failure, with a case-fatality rate of 11% to 25% (Marston et al., 1994; Stout & Yu, 1997). Pontiac fever is milder and characterized by fever, fatigue, myalgia, and headache, with or without cough (Tossa et al., 2006), and patients recover within two to five days without treatment. Individuals at increased risk of legionellosis include older adults, males, smokers, and those with underlying conditions, including immunosuppression, chronic lung disease, diabetes, and cancer (Marston et al., 1994; Stout & Yu, 1997).
Legionella bacteria occur naturally in freshwater and soil and multiply in environmental protozoa. They become a human health risk when they multiply in water between 25°C and 42°C, become aerosolized, and are inhaled into the lungs (Fields et al., 2002; WHO, 2007). Infection with Legionella pneumophila (which causes more than 70% of human infections) occurs through inhalation of aerosols from manufactured freshwater systems in homes or in the community (e.g., cooling towers, hot tubs, shower heads, fountains) (WHO, 2007). Transmission also occurs in care facilities due to a combination of inadequate design and/or maintenance of building infrastructure and risk factors present in the residents or patients (Silk et al., 2013). Infection with some species occurs through direct contact with, or via aerosolization of, contaminated soil or compost (WHO, 2007; Picard-Masson et al., 2016).
As an environmental bacterium, Legionella is sensitive to climatic factors. Legionellosis shows a marked seasonality, with higher incidence in the late summer and early fall (Simmering et al., 2017; Alarcon Falconi et al., 2018). It also shows regional differences, which may be associated with climatic factors (Beauté et al., 2016; Simmering et al., 2017). The most consistent and strongest climate association has been reported between legionellosis and precipitation. Several studies in North America, Asia, and Europe have found an increase in the risk of sporadic legionellosis following rainfall events (Fisman et al., 2005; Hicks et al., 2007; Garcia-Vidal et al., 2013). The increase occurs two to 11 days after rainfall (Fisman et al., 2005; Chen et al., 2014b; Beauté et al., 2016;) and has been associated with a 2.5 to 2.6-fold increase in incidence per 5 mm to 1 cm of rain (Hicks et al., 2007; Chen et al., 2014b) or 2.1 to 2.5-fold compared to periods with no rain (Fisman et al., 2005; Beauté et al., 2016). This may be because precipitation increases runoff and adds nutrients and protozoa to water sources, which can increase Legionella replication; it can also increase turbidity and organic matter, which decreases the effectiveness of water disinfection (Fisman et al., 2005).
In temperate zones, warmer temperatures have also been found to increase the risk of legionellosis (Fisman et al., 2005; Hicks et al., 2007; Conza et al., 2013; Halsby et al., 2014; Beauté et al., 2016; Simmering et al., 2017). An increase of 1°C has been associated with a 2.8% to 7% increase in risk (Fisman et al., 2005; Hicks et al., 2007; Conza et al., 2013), and the risk occurs after a lag of three to nine weeks (Halsby et al., 2014; Beauté et al., 2016). However, one study (Beauté et al., 2016) suggested that risk may decrease at higher temperature ranges (above 20°C). Finally, higher relative humidity and vapour pressure have been associated with legionellosis risk (Fisman et al., 2005; Conza et al., 2013; Gleason et al., 2016; Simmering et al., 2017). Higher humidity has been positively associated with legionellosis cases (Gleason et al., 2016). Simmering et al. (2017) found that temperature and humidity interact; the highest risk was between 16°C and 27°C and a relative humidity of more than 70%. In summary, the risk of legionellosis seems to increase following warmer, wetter weather (Chen et al., 2014a; Halsby et al., 2014), as warm, wet environments support Legionella growth and aerosolization.
6.3.4.2 Cryptococcus gattii Infection
Cryptococcus gattii is an environmental fungus that causes infection in humans and multiple animal species (Chen et al., 2014b). Cryptococcus gattii occurs in soil and tree debris, but it can also be found in air, freshwater and saltwater, and on vehicles and fomites — objects or materials such as clothes, utensils, and furniture that can carry infection (Kidd et al., 2007a). Its spores are aerosolized from soil or trees and transported to other sites or inhaled by humans and animals (Kidd et al., 2007b). In humans, spores are inhaled into the lungs, where they cause pneumonia or pulmonary nodules. They may be disseminated through the bloodstream, particularly to the central nervous system, where they cause meningitis or brain nodules (Chen et al., 2014b) (Figure 6.7). The case-fatality rate is high, at 23% (Phillips et al., 2015). Individuals at higher risk of infection include those over 50 years of age, smokers, and people with chronic lung disease or immunocompromizing conditions or treatments (MacDougall et al., 2011).
Cryptococcus gatti lives as a yeast in the environment (1), usually in association with certain trees or soil around trees. Humans and animals can become infected with C. gattii after inhaling airborne, dehydrated yeast cells or spores (2), which travel through the respiratory tract and enter the lungs of the host (3). The small size of the yeast and/or spores allows them to become lodged deep in the lung tissue. The environment inside the host body signals C. gattii to transform into its yeast form, and the cells grow thick capsules to protect themselves (4). The yeasts then divide and multiply by budding. After infecting the lungs, C. gattii cells can travel through the bloodstream (5) — either on their own or within macrophage cells — to infect other areas of the body, typically the central nervous system (6).
Source
US CDC, 2020.
Cryptococcus gattii is found worldwide. It first emerged in Canada on Vancouver Island in 1999, when it caused an outbreak among healthy animals and humans (Fyfe et al., 2008). Since then, incidence increased to reach a steady state of 0.2 to 0.5 cases per 100,000 population in British Columbia (BCCDC, 2019). Since 2004, the fungus has been found in the environment and in humans and animals on the British Columbia mainland and neighbouring Washington and Oregon states (MacDougall et al., 2007). It is unclear whether the range of C. gattii increased or whether C. gattii was already present in these regions (Roe et al., 2018). Very few locally acquired Canadian cases have been reported outside British Columbia, except for two human cases in Quebec in 2008 and 2015 (St-Pierre et al., 2018) and one case in a deer in Nova Scotia in 2014 (Overy et al., 2016).
As an environmental fungus, C. gattii is sensitive to climatic factors. Humidity, temperature, precipitation, evaporation, and solar radiation all affect the occurrence of C. gattii in the environment (Granados & Castañeda, 2006; Kidd et al., 2007b). An ecological niche model conducted in British Columbia described areas with environmental conditions supporting C. gattii establishment as those with low elevations, with a daily January average temperature of above 0°C, and within two coastal climatic zones of British Columbia, where winters are mild and wet and summers are warm and dry (Mak et al., 2010). Environmental sampling of C. gattii in British Columbia found that air sample concentrations were greatest during summer months with warm, dry conditions, and low relative humidity (Kidd et al., 2007b).
The response to climatic and other environmental factors has been inconsistent globally and may depend, in part, on where C. gattii is sampled (i.e., air, soil, or trees), on strain type, and on other factors. Wind increases aerosolization from the soil and enables spread (Uejio et al., 2015). Heavy rainfall or humidity may pull spores out of the air (Kidd et al., 2007b), although in most regions, C. gattii is more likely to be isolated from trees during the rainy season, suggesting it is more abundant at this time (Uejio et al., 2015). In British Columbia, higher temperatures decreased isolation from trees and concentration in soil, but in other parts of the world, the highest isolation frequency was during very high temperatures (Uejio et al., 2015). Cryptococcus gattii spores can withstand high levels of solar radiation, so summer conditions with high temperatures and high isolation are unlikely to preclude C. gattii survival (Rosas & Casadevall, 2006).
6.3.4.3 Blastomycosis
Blastomyces dermatitidis is a dimorphic fungus found in moist soil and decaying vegetation. Transmission occurs from the environment to humans and other animals through aerosolization and inhalation (Castillo et al., 2016). Spores inhaled into the lungs cause pulmonary blastomycosis, characterized by cough, fever, and shortness of breath (McBride et al., 2017). The yeast form may be disseminated through the blood to other organs, and cutaneous infection can occur from direct inoculation (Castillo et al., 2016). The case-fatality rate is high, at 5% to 20%, which may be associated with delayed diagnosis, particularly in less endemic areas (Crampton et al., 2002; Dalcin & Ahmed, 2015). The infection is treated with antifungals (Castillo et al., 2016). The incubation period is typically one to three months and cases mainly occur in immunocompromised individuals, smokers, and those with underlying diseases who are at higher risk of infection and mortality (Crampton et al., 2002; Dalcin & Ahmed, 2015; Castillo et al., 2016; McBride et al., 2017). Individuals are at increased risk of exposure through outdoor occupations and activities such as hunting and camping (Crampton et al., 2002). Most cases are sporadic, but activities that disrupt soil (e.g., construction, deforestation, outdoor activities) and exposure to riverbanks have been associated with point-source outbreaks (Baumgardner & Burdick, 1991; Proctor et al., 2002; Azar et al., 2015).
Blastomyces dermatitidis is endemic in certain parts of Canada, including Southern Saskatchewan, Manitoba, Ontario, and Southern Quebec (Crampton et al., 2002; Litvinov et al., 2013; Lohrenz et al., 2018). Blastomycosis is only reportable in Ontario and Manitoba and may be under-recognized in other parts of the country. It is hyperendemic in Northwest Ontario, characterized by persistent, high levels of disease occurrence. The Kenora, Ontario, area has the highest reported rates worldwide, with 35 hospitalizations per 100,000 population per year (Litvinjenko & Lunny, 2017; Brown et al., 2018). This endemic region is contiguous with endemic regions in US states that border the Mississippi and Ohio rivers, the Great Lakes, and St. Lawrence Seaway (McBride et al., 2017).
Blastomyces species are environmental fungi and therefore depend on climatic conditions for growth and dispersal. Specific ecological factors are poorly understood, in part because the organism has rarely been recovered from the environment, but moist soil near waterways and lakes seem to be the preferred niche for Blastomyces species fungi (Castillo et al., 2016). Soil disruption facilitates aerosolization (Baumgardner, 1997). It is hypothesized that proximity to waterways and drought followed by increased precipitation may help expose spores and facilitate their dispersal (McDonough et al., 1976; McTaggart et al., 2016). Although the peak onset for pulmonary infection is in the fall and winter, reflecting exposure during warmer weather (Morris et al., 2006; Light et al., 2008; Dalcin & Ahmed, 2015; Brown et al., 2018), few studies have specifically assessed the role of climatic factors and varying results have been reported. Baumgardner et al. (2011) found an association between total precipitation, temperature (low average temperatures during the season of infection, but high maximum temperatures the previous season), and canine cases. Seitz et al. (2015) found a higher risk of blastomycosis hospitalization with lower maximum temperature. Proctor et al. (2002) investigated an outbreak that followed a prolonged period of drought. Similarly, Pfister et al. (2011) found that moderate-to-severe drought, followed by above-normal precipitation and warming of yard waste, was associated with an outbreak in Wisconsin. Wind may be a factor, as some outbreaks have been associated with excavation dust (Baumgardner et al., 2011) or disruption of decomposing yard waste (Pfister et al., 2011). A Wisconsin ecological niche model found proximity to waterways and a summer vegetation index to be most predictive of the location of human and animal blastomycosis cases; bioclimatic factors did not play a significant role (Reed et al., 2008).
6.3.4.4 Histoplasmosis
Histoplasmosis is caused by the dimorphic fungus Histoplasma capsulatum. Spores grow in moist soil rich in bat or bird guano along waterways and in caves. Disruption of soil due to excavation and construction leads to aerosol generation and the risk of inhalation of spores. Infection develops seven to 14 days after exposure. It is often asymptomatic or causes self-limited illness with fever, headache, weakness, chest pain, and cough. It can lead to extrapulmonary infection and also cause more severe pulmonary infection in individuals with underlying lung conditions or immunocompromised individuals (Kauffman, 2007). Histoplasma capsulatum is endemic along the St. Lawrence and Great Lakes River Drainage Basins in Quebec and Ontario, the Ohio and Mississippi River valleys in the United States, and along waterways in Asia and South America. In Canada, the highest rate of diagnosed illness is in Quebec. In one study, 58 cases were identified as having occurred in Canada over three years (1992 to 1994), of which 72% were from Quebec (Nicolle et al., 1998). Cases in Alberta were first identified in 2003, and since then a small number are reported every year (Anderson et al., 2006; Alberta Health, 2018). The fungus is climatically sensitive, preferring moderate temperatures (18°C to 28°C), constant humidity, and low light (Teixeira et al., 2016).
6.3.4.5 Coccidioidomycosis
Coccidioidomycosis is caused by Coccidioides species fungi, which are found in arid soil, including C. immitis found in California and C. posadasii found elsewhere (Maves & Crum-Cianflone, 2012). Spores are released into the air when soil is disturbed and can be inhaled by humans, causing coccidioidomycosis, also known as valley fever. The disease has an incubation period of one to three weeks, and many individuals remain asymptomatic or have mild symptoms. Approximately 40% develop pulmonary infection with cough, chest pain, dyspnea, and fever. This is usually self-limited and resolves within a few weeks. Disseminated infection occurs in 5% of individuals and leads to diffuse and chronic pulmonary illness, meningitis, or infection in other organs (Maves & Crum-Cianflone, 2012). This severe form requires prolonged antifungal treatment. Pregnant women and people with impaired cellular immunity, due to HIV infection or organ transplantation, are at higher risk of severe illness. Reactivation of latent disease is also possible (Maves & Crum-Cianflone, 2012). The geographic range of the fungus includes parts of South and Central America, Mexico, and the Southwestern United States (Maves & Crum-Cianflone, 2012). Certain areas of California and Arizona have the highest reported rates worldwide. Soil disruption, including construction, archaeological digs, earthquakes, and wind storms in endemic desert areas is the main source of exposure (Maves & Crum-Cianflone, 2012).
Coccidioides spp. are affected by climatic factors (Nguyen et al., 2013), requiring moisture to grow in soil and then a dry period to mature into spores that can be aerosolized (the “grow and blow” hypothesis) (Comrie & Glueck, 2007). The seasonal spike in human cases in the United States occurs mainly in the fall, after a period of rainy months followed by warm dry months. There is also a cyclical pattern, with incidence peaks approximately every two to five years, believed to be associated with droughts followed by rainy weather (Park et al., 2005; Tamerius & Comrie, 2011; Gorris et al., 2018 ). Warmer temperatures are associated with higher incidence in humans (Gorris et al., 2018), and incidence increases following large windstorms (Tong et al., 2017). However, regions with high mean annual precipitation or very moist soil (such as coastal regions) have a low incidence (Gorris et al., 2018).
6.4
Adaptation to Reduce Health Risks
Share
Canada already has a robust public health system, which is networked with international public health organizations. Together, these systems contribute to infectious disease preparedness, surveillance, and monitoring and outbreak response. As infectious disease risks continue to grow in a changing climate, these activities will be increasingly important for adaptation success. Two streams of public health activities in the context of emerging and re-emerging infectious diseases are upstream preparedness and outbreak response. Within these streams, there are three key public health activities: risk assessment, to identify current and future risk; surveillance, for known risks or possible future emerging risks identified in risk assessments; and interventions, to prevent and control infectious diseases. These three activities are discussed individually, but, in practice, they should be highly connected; risk assessments guide rational implementation of surveillance activities, which, in turn, trigger interventions to protect the public when a hazard is detected (Figure 6.8). Actions to protect health need to be robust and coordinated to address the increased risks from infectious diseases due to climate change. Furthermore, as many of the threats of emerging and re-emerging infectious diseases involve directly transmitted or vector-borne zoonoses, a One Health approach, which considers the interactions among human, animal, and environmental factors in disease transmission, emergence, and re-emergence, is required to develop effective responses (Ogden et al., 2017; Ogden et al., 2019).
In general, Canada and other high-income countries could be considered well-placed to respond to emerging and re-emerging infectious diseases through robust international institutions for surveillance and response, such as the WHO’s International Health Regulations (IHR) (WHO, 2019c), the Global Outbreak Alert and Response Network (WHO, 2019d), as well as the health care system and public health institutions in Canada. The public health system in Canada centres around the Pan-Canadian Public Health Network (Pan-Canadian Public Health Network, 2016), a federal, provincial, and territorial partnership, with federal coordination by PHAC. This system is supported by high-quality academic institutions and industries in Canada that can facilitate innovation and responses to threats from emerging infectious diseases. Canada is a leader in the One Health approach in federal, provincial, and academic institutions, and the One Health approach is applied systematically in addressing these disease threats (Government of Canada, 2013; INSPQ, 2018a; CPHAZ, 2019; GREZOSP, 2019; Ogden et al., 2019). Despite this capacity, there are many unknowns ahead, and, as seen with COVID-19, even the best-prepared public health systems can be severely challenged by emerging infectious diseases.
Two streams of public health actions to manage emerging and re-emerging infectious diseases.
Figure 6.8
The blue boxes indicate outbreak response, and the orange boxes indicate upstream preparedness. Examples of activities that may be components of outbreak management are shown in the green boxes. Dashed lines indicate how outbreak response may be triggered by early detection of disease risk using an upstream preparedness approach.
Since the last national Canadian climate change and health assessment (Berry et al., 2014a), PHAC enhanced upstream preparedness for emerging and re-emerging vector-borne disease risks through an integrated program of adaptation, which comprises (Figure 6.8):
prioritization of disease risks for study, now formalized in a multi-criteria decision analysis (MCDA) method (Ng et al., 2019);
modelling studies to identify climate-disease risk associations and to project where and when diseases may emerge with climate change (Ng et al., 2019);
implementation of surveillance programs to validate models and track emerging vector-borne diseases (Drebot, 2015; Ogden et al., 2014c); and
development of synthesized knowledge (Hierlihy et al., 2019), disease forecasting (Ripoche et al., 2019), and risk communication tools (Figure 6.9) to support local efforts to adapt to increasing risks from vector-borne diseases.
6.4.1
Risk Assessment to Identify Current and Future Risks
Share
6.4.1.1 Responsive Rapid Risk Assessments
Responsive rapid risk assessments (RRAs) are qualitative risk assessments developed in response to an immediate disease emergence event within Canada, or a perceived risk, such as an international disease event or threat, that requires rapid decision making. Responsive RRAs are developed using the scientific literature and knowledge of a disease at the moment in time that the disease emerges or re-emerges; they may be updated as knowledge increases in the light of the disease emergence event. They generally pre-date, and aim to guide, implementation of surveillance and/or intervention activities. Examples include those conducted in response to the threat of Zika virus in Canada nationally (Government of Canada, 2016) and provincially (INSPQ, 2016), as well as those developed by the European Centre for Disease Prevention and Control (ECDC) in response to a number of events (ECDC, 2019). The World Health Organization has developed a protocol for their preparation (WHO, 2019e), which forms the basis for RRAs used in Canada. The capacity to perform RRAs in the face of disease emergence events is essential for public health officials in Canada to respond to unforeseen events, including those driven by climate change.
6.4.1.2 Predicting Future Populations at Risk, and Establishing Early Warning Systems, Through Quantitative Model-Based Risk Assessments
Quantitative risk assessments aim to predict where and when infectious diseases may emerge or re-emerge with a precision useful to public health officials in their efforts to prepare and respond to disease emergence with needed policies or programs. Risk assessments can be obtained using mathematical models that recreate the essential components of vector life cycles or disease transmission cycles, if there are sufficient quantitative data to calibrate these types of models. If not, statistical or ecological niche models can be used, providing there are surveillance data that include, at a minimum, information on presence, but preferably data on presence/absence and abundance, to calibrate them. In either case, the objective of the models is to assess where and when risk from climate-sensitive infectious diseases may change (i.e., they provide “early warning” of future disease risks). Predictive models are used for three main purposes (Ogden & Gachon, 2019):
short-range forecasting of disease risks based on weather;
long-range or seasonal forecasting based on climate; and
projections of long-term effects of climate change on disease risks.
Models are developed by first identifying key associations between the presence of vectors and/or pathogens and weather (for short-term forecasting) or climatic variables, while accounting for important climate-independent determinants of their presence, which may include other environmental changes such as land-use change (Patz et al., 2003). Validation of deduced associations against real data is important to provide confidence in the use of model outputs. Once associations between climate and the occurrence of pathogens/vectors have been identified and quantified, the impacts of projected climate change, obtained from regional and global climate models, on pathogen/vector distributions and risk can be estimated.
Forecasting provides early warning of impending weather-driven disease outbreaks. This can include short-range forecasting on a time scale of days to weeks and long-range forecasting on a time scale of several months that can be used to implement prevention and control activities (Morin et al., 2018). The model-based assessments of the effects of climate change provide assessments of where and when diseases may emerge or re-emerge in coming decades, which are used in a number of contexts (Ogden et al., 2014c):
high-level national policy decisions on whether and when public health preparedness for emerging risks may be needed (Centre for Food-borne, Environmental and Zoonotic Infectious Diseases, 2017);
provincial, territorial, and municipal assessments of vulnerability to the health impacts of climate change and of needed adaptation measures (Berry et al., 2014b); and
design and implementation of surveillance (Ogden et al., 2014).
As discussed earlier in this chapter, a number of model-based risk assessments have been conducted for Canada, including those for Lyme disease (Ogden et al., 2006; Ogden et al., 2014), relapsing fever (Sage et al., 2017), Dermacentor variabilis ticks (Minigan et al., 2018), exotic mosquito-borne diseases and their vectors (Ogden et al., 2014a; Ng et al., 2017), and WNV (Hongoh et al., 2012; Chen et al., 2013).
6.4.2
Prioritization
Share
Since they were identified as an important methodological need by Charron et al. (2008) and by the WHO (WHO, 2019g), a number of MCDA methods have been explored to prioritize diseases for attention, based upon their public health importance, including their risk of emergence or re-emergence due to climate change (Ng & Sargeant, 2012; Cox et al., 2013). These types of methods have been adopted and occasionally adapted for disease prioritization at both federal (Otten et al., 2019) and provincial levels (INSPQ, 2018b), although not as yet by all jurisdictions in Canada.
6.4.3
Surveillance for Known Risks or Possible Future Risks
Share
There are two types of surveillance to identify emerging infectious diseases to inform outbreak management. The first is vigilance for international disease emergence events, and the second is disease-specific surveillance, to be implemented once a disease is recognized as a significant public health risk for Canadians. The possible impact of climate change on increasing infectious disease emergence and re-emergence events globally, and on increasing dispersion of the disease agents through travel, trade, and migration, increases the need for effective vigilance for emerging infectious disease events in other countries. International efforts that support this type of surveillance include the IHR, under which all WHO member states are obliged to undertake surveillance for disease emergence events and to report any that are identified. In addition, as part of the global One Health initiative, the Food and Agriculture Organization, World Organization for Animal Health, and the WHO collaborate in the Global Early Warning System, which is a warning system for emerging disease threats at the human–animal–ecosystems interface. International “passive” surveillance programs include the GeoSentinel Surveillance Network, an international network of voluntarily participating medical clinics, and surveillance systems that scrutinize publicly available web content for signals of potential emerging threats. These include the Program for Monitoring Emerging Diseases (ProMed), HealthMap, and MediSys, as well as Canada’s own Global Public Health Intelligence Network (GPHIN), operated by PHAC (Ogden et al., 2017).
In Canada, a number of methods are available and used for the surveillance of endemic and emerging infectious diseases, to follow trends in incidence, identify the spatiotemporal distribution and spread of cases (including clusters) and risk, and identify risk factors in the affected population. The standard method for acquiring data on human disease cases nationally is through the National Notifiable Disease Surveillance System, coordinated by PHAC (Government of Canada, 2019e). Some diseases that are not nationally notifiable may be notifiable at provincial and territorial levels. These notifiable diseases are identified using clinical and laboratory case definition criteria, and emergence or re-emergence (nationally or internationally) is one criterion. Other criteria for adding diseases to the list include diseases under international surveillance, considerations of incidence in Canada, severity, communicability, potential for outbreaks, socio-economic burden, preventability, risk perception, and necessity for public health response (Public Health Agency of Canada, 2009).
Other methods of capturing information on possible human disease cases during emergence events include syndromic surveillance; for example, the use of real-time data from pharmacies on sales of influenza remedies were used to track the evolution of pandemic H1N1 influenza in Canada in 2009 (Muchaal et al., 2015). Another method is passive laboratory-based surveillance. In response to possible increasing risks from arboviruses in Canada, PHAC’s National Microbiology Laboratory instigated surveillance for human cases of non-WNV arboviruses by routinely taking sera submitted for WNV testing that were WNV-negative, and testing them for a panel of endemic and exotic arboviruses (Drebot, 2015). The Pan-Canadian Public Health Network has the capacity to instigate national human case surveillance for limited periods during disease emergence events. It recently used this capacity in response to the COVID-19 pandemic and to the the Zika outbreak in Central and South America in 2015–2016, which infected more than 500 Canadian travellers (Government of Canada, 2019f).
There has been much effort in Canada to use surveillance in sentinel animals and vectors to detect risks from emerging vector-borne zoonoses, particularly WNV, Lyme disease, and other Ixodes species-transmitted pathogens, as a method of early warning of the spread of emerging diseases, and, for WNV, of impending outbreaks (Thomas-Bachli et al., 2015). For vector-borne diseases, vector surveillance includes active surveillance to capture vector mosquitoes, which are then tested for mosquito-borne pathogens, particularly WNV, which is carried out routinely in many jurisdictions in Canada (Government of Canada, 2019a). It also includes the collection of ticks by drag sampling, the capture of tick hosts, and the testing of ticks and hosts for tick-borne pathogens (Ogden et al., 2014). Passive surveillance for ticks, involving submission of ticks from the patients of medical and veterinary clinics, has been a key surveillance activity in Canada, providing a uniquely long dataset that has supported attribution of I. scapularis range expansion in Canada to impacts of climate change (Leighton et al., 2012).
For both mosquitoes and ticks, the surveillance for vectors is capable of capturing multiple species of vectors and pathogens. Mosquito surveillance targeting WNV has identified incursions of exotic Aedes spp. vectors in Southern Ontario, which are vectors of dengue virus and Zika virus (Windsor-Essex County Health Unit, 2019a, Windsor-Essex County Health Unit, 2019b). Tick surveillance is undertaken predominantly by two methods: passive tick surveillance, involving submission of ticks that members of the public found on themselves or their pets via participating veterinary and medical practices; and active tick surveillance, through collecting host-seeking ticks by dragging a flannel across the ground (drag sampling), or through examining captured wild rodents (Ogden et al., 2014c). These forms of tick surveillance were originally designed to identify risk from Lyme disease, but can detect risk from other species of tick vectors of disease (Gabriele-Rivet et al., 2015; Nelder et al., 2019). Ticks from both types of surveillance are routinely tested for a wide range of pathogens, in addition to the bacteria causing Lyme disease (Dibernardo et al., 2014). In Canada, most ticks collected through surveillance are currently identified to species in provincial laboratories, and then tested for pathogens at the National Microbiology Laboratory. However, this method will not be sustainable once ticks become more widespread, given the expected and observed (Gasmi et al., 2018) large increase in the number of tick submissions across Canada. New and less resource-intensive surveillance systems are needed to protect Canadians from the increasing risks of these diseases.
A citizen science project called eTick was launched in 2014. It is a web-based platform that allows members of the public to submit images of ticks for species identification by an entomologist (eTick, 2019). It greatly expands the potential geographical coverage of tick monitoring, in addition to providing an opportunity to deliver timely public health information directly to the population exposed to ticks. It allows for the real-time monitoring and mapping of various tick species, including changes in distribution. The program is currently available in Quebec, Ontario, and New Brunswick. In Manitoba, an image-identification system called the Tick Checker, which is similar to eTick, was implemented in 2017 (Government of Manitoba, 2019a). This effort is part of Manitoba’s black-legged tick passive surveillance program, which identifies locations where new tick populations and Lyme disease risk may be emerging, before confirmation by active tick surveillance (Government of Manitoba, 2018).
Livestock health organizations, such as the Canadian Food Inspection Agency, track some zoonoses in sentinel animals (for example, EEEV in horses) when they fall within the mandate of these organizations (Government of Canada, 2018c). Wildlife health organizations, such as the Canadian Wildlife Health Cooperative, assist in tracking some wildlife-borne zoonoses in sentinel animals (CWHC, 2019).
6.4.4
Interventions to Prevent and Control Disease Incidence
Share
International responses to emerging infectious disease outbreaks are mandated under the IHR through the Global Outbreak Alert and Response Network (GOARN) (WHO, 2019d). This network brings together technical and operational resources from relevant institutions in WHO member states, including surveillance initiatives, networks of laboratories, United Nations organizations (e.g., UNICEF, UNHCR), the Red Cross, and international humanitarian non-governmental organizations, such as Médecins sans frontières. The network has the capacity to assemble and deploy technical teams to countries affected by outbreaks to assist with coordination, clinical disease management, epidemiological analysis, as well as logistics and communications that are needed to control outbreaks. PHAC is the lead for Canada’s extensive participation in GOARN. The agency was created in 2004, in part to ensure and enhance national coordination of responses to emerging infectious diseases through the Pan-Canadian Public Health Network and to participate in the IHR and GOARN.
A key capacity for reducing risks to Canadians from outbreaks is the health care system and its ability to identify human disease cases and treat them. Diagnosis and treatment of cases reduces transmission and contributes to controlling outbreaks. However, this requires vigilance by front-line health care workers; in some outbreaks, health care facilities can become a focus for transmission, as occurred during the SARS outbreak in Canada (Varia et al., 2003). Increasing antimicrobial and antiviral drug resistance challenge the capacity of health care systems to treat emerging diseases (WHO, 2019f).
Canada’s technological capacity to prevent and control infectious diseases is well developed. The Federal/Provincial/Territorial Canadian Public Health Laboratory Network, supported by PHAC’s National Microbiology Laboratory, which is one of only seven level-4 containment facilities in North America, has the capacity to develop, improve, and implement diagnostic tests for emerging infectious diseases such as Zika virus (Safronetz et al., 2017). The global network of public health organizations and pharmaceutical industries are capable of developing vaccines in response to outbreaks (Carlsen & Glenton, 2016; Henao-Restrepo et al., 2017). The WHO has developed a list of priority pathogens for which vaccines are urgently needed; several of these diseases are zoonoses with the potential for person-to-person spread, and emergence risk may be enhanced by climate change. However, the time it takes to develop vaccines, and to assess them for safety and efficacy to ensure their public acceptability (Carlsen & Glenton, 2016), emphasizes the need for upstream preparedness rather than simply relying on vaccine development to manage outbreaks. While there is much capacity for vaccine discovery in Canada (Plummer & Jones, 2017), the COVID-19 pandemic has highlighted the lack of Canadian capacity for vaccine manufacture.
Other technological adaptations are important for current and future vector-borne disease control in Canada. For mosquito-borne diseases, such as WNV, mosquito control methods are effective mainstay methods; these include killing larvae in their aquatic habitat (larviciding) using non-chemical biological control agents such Bacillus thuringiensis israelensis, and killing adult mosquitoes (adulticiding) using chemical sprays such as deltamethrin (Reisen & Brault, 2007). For tick-borne diseases, such as Lyme disease, vector control methods, such as area-wide spraying with acaricides, remain unlicensed, and thus unavailable, or at an experimental stage of development in Canada (Ogden et al., 2015). Personal protection measures remain the most effective of control of vector-borne diseases, and these include insect repellents, which prevent tick bites, insecticide (permethrin)-treated clothing, and prompt removal of feeding ticks from the body to prevent transmission of B. burgdorferi (Ogden et al., 2015). For these measures to be effective, the public needs to understand the risks, perceive them to be worth acting on, know what protective actions to take, and be willing to take them. In turn, this requires effective communications from public health organizations at all levels, which is an increasingly difficult task because misinformation, particularly from the internet, is an increasing problem (Greenberg et al., 2017). As well as communicating information about personal disease protection in Canada, information for travellers will become increasingly important. This will help prevent them from introducing infections upon returning to Canada, particularly exotic vector-borne diseases, which may be increasingly likely to be autochthonously transmitted in this country under climate change (Berrang-Ford et al., 2009).
6.4.5
Adaptation Challenges for Communities and Members of Society at Increased Risk
Share
Vulnerability to infectious disease health risks related to climate change is complex and determined by a number of factors that interact to influence an individual’s exposure, sensitivity, and adaptive capacity. While all Canadians are vulnerable to climate change, the experiences of impacts and risks are not uniform, and some individuals and communities will be disproportionately affected (see Chapter 9: Climate Change and Health Equity). Existing health inequities, and variations in the status of determinants of health, in combination with exposure (e.g., geographic location), and genetic and biological factors, can increase or decrease climate change vulnerability. There are existing knowledge gaps regarding the combination of variables that enhance vulnerability to infectious disease risks related to climate change. However, it is expected that individuals with determinants of ill health (e.g., low-income, living in substandard housing, food-insecure) are more likely to experience disproportionate impacts and have limited ability to take protective measures. For example, in general, poorer health status is associated with lower socio-economic status, which, in turn, is associated with higher rates of unhealthy behaviours and dietary habits (Hajizadeh et al., 2016) that lead to higher rates of chronic diseases (Roberts et al., 2015) and can increase health risks associated with infectious diseases related to climate change.
In Canada, First Nations, Inuit, and Métis peoples experience lasting, systemic health inequities, which are associated with a legacy of colonization and intergenerational trauma (PHAC, 2019). They experience significantly higher rates of infectious diseases compared to non-Indigenous populations, which increases their risk of climate change-related infectious diseases (Adekoya et al., 2015). While greater sensitivity to infectious diseases, which may be due to genetic factors or high rates of chronic co-morbidities (Badawi et al., 2018), have been observed (e.g., genetic susceptibility to H1N1 influenza), existing health inequities, such as limited availability and accessibility of acceptable health care services, overcrowded housing, and lack of culturally relevant disease-prevention information (National Collaborating Centre for Aboriginal Health, 2016) can compound vulnerability (see Chapter 9: Climate Change and Health Equity). In addition, First Nations, Inuit, and Métis peoples may be at increased risk of exposure due to a strong reliance on traditionally harvested foods and land-based activities, such as hunting. For example, outbreaks of trichinellosis have been associated with the consumption of marine mammals in Arctic regions (Yansouni et al., 2016), and a rise in the abundance of mosquitoes, ticks, and other biting insects that have the potential to transmit new vector-borne diseases is of increasing concern (see Chapter 2: Climate Change and Indigenous Peoples’ Health in Canada). Geographic location may also increase exposure; for example, risk of exposure to Arctic fox rabies is higher for Arctic communities.
Policies to address these underlying causes of vulnerability are generally not within the mandate of public health organizations (Adler & Newman, 2002); however, there is increasing recognition in the public health field that climate change will exacerbate health inequities (Friel, 2019). Efforts to strengthen determinants of health should be combined with climate change and health activities (e.g., adaptation measures) (see Chapter 9: Climate Change and Health Equity).
6.4.6
Adaptation Gaps and Opportunities
Share
6.4.6.1 Adaptation Gaps
Prioritizing adaptation options to manage infectious disease risks from climate change in Canada should take into consideration multiple criteria, including the immediacy and level of the risks, the technical viability of the options, human and financial resources, compatibility with current policies, and other constraints (Ebi et al., 2006a; Ebi et al., 2006b). For many of the infectious disease risks associated with climate change discussed in this chapter, surveillance, monitoring, and disease control will only be successful with local action, so enabling local public health and health care systems to undertake these activities will be important in enhancing resilience to these threats. In a review of national-level adaptation planning for climate change impacts on infectious diseases in 14 Organization for Economic Co-operation and Development member nations, a number of limitations to current planning were observed. These included negligible consideration of the needs of population groups that experience disproportionate impacts, limited engagement at local government levels to assess health risks, and inadequate logistics and support related to funding, timelines for assessments, and development of adaptation plans (Panic & Ford, 2013). The report highlighted that these limitations need to be addressed in Canadian adaptation plans. Post-pandemic evaluation of Canadian public health responses to COVID-19 will likely be highly informative of the current adaptive capacity gaps.
In addition, adaptation to climate-related infectious diseases is constrained by existing research gaps. For many climate-related infectious diseases, there is a lack of knowledge of their ecology, little surveillance data for calibration of statistical/ecological niche models and for validation of predictive models, and a lack of long-term monitoring that is required to attribute changes in disease risk to climate change (Ebi et al., 2017). Effective control of tick-borne diseases in the environment is particularly difficult, and research is needed in this field. Knowledge and research gaps are being identified using scoping and systematic reviews about specific vector-borne disease risks prioritized by MCDA methods (Otten et al., 2019).
A significant challenge in protecting the health of Canadians is that, globally, the capacity to control vectors is diminishing because of increasing resistance against insecticides. However, techniques such as the introduction of genetically modified mosquitoes that control populations by producing non-viable offspring, and mosquitoes carrying Wolbachia bacteria in their guts, rendering them incompetent as vectors, are a focus of research. These techniques may become available to public health officials in the near future.
6.4.6.2 Adaptation Opportunities
Existing information on the ecology and epidemiology of vector-borne diseases that threaten Canadians (Corrin et al., 2018), as well as prevention and control methods and their efficacy (Hierlihy et al., 2019), are being provided to public health officials at all levels of government to assist in developing response plans. The expected increased epidemic behaviour of mosquito-borne diseases such as WNV and EEEV means that weather-based forecasting of disease risk, as recommended by the IPCC, would be helpful for use in early warning systems. Much progress has been made to develop methods for weather-based forecasting for WNV, but implementation in Canadian public health programs will require further work (Ogden et al., 2019).
The increasing possibility of autochthonous transmission of exotic diseases poses a challenge for public health activities and the Canadian health care system. Since exotic diseases have been absent in Canada, health professionals have screened them out as possible diagnoses in patients without a history of travel to well-known endemic areas. An increase in modelling of disease risks will enhance preparedness for disease emergence events, and greater surveillance of new non-endemic infectious diseases among patients seeking care would be prudent.
Similarly, enhanced surveillance is required for vector-borne diseases that are currently exotic to North America but could be carried into Canada by travellers, those that are absent from Canada but could expand northward into Canada from the United States, and those that are Canada-endemic and may re-emerge in the form of outbreaks. Education of health care providers about these possible new emerging diseases in Canada will be important for early diagnosis and treatment from the perspective of affected individuals, but also for early detection of outbreaks from a public health perspective. Citizen-based approaches to surveillance, such as the eTick program (eTick, 2019), are also an increasingly recognized opportunity. These approaches could also be enhanced by Indigenous knowledge and observation (Tomaselli et al., 2017; Henri et al., 2018).
There are opportunities to employ “big data” and technologies such as artificial intelligence (AI) that help analyze big data to address growing risks to health from infectious diseases. The first big data source that is being used in this regard is modern genomics. Whole genome sequencing (WGS) is now mainstreamed in Canadian enteric disease surveillance through PulseNet, as it is in the United States and Europe. PulseNet allows for more sensitive detection of disease clusters and outbreaks and greater capacity to reliably attribute sources of infection (Gilmour et al., 2013). The immense size of the data generated by WGS means that complex bioinformatics analysis methods have had to be developed for timely analysis of the data (IRIDA, 2019). Application of WGS to other applications in infectious disease diagnosis and surveillance, and further development of molecular epidemiology to make full use of the data, will continue and will support more robust efforts to tackle these risks. Metagenomic and meta-barcoding methods, which allow the identification of the range of species present in complex mixed-species samples, lend themselves to widened surveillance of mosquitoes and mosquito-borne diseases, including exotic mosquitoes and pathogens. Through these methods, the range of mosquito species and mosquito-borne pathogens, rather than simply WNV, is identified in mosquito-trap captures obtained during mosquito surveillance. These methods may also revolutionize the detection of pathogens in other environmental DNA samples from drinking and recreational water and from soil.
The second big data source is web media and social media, obtained by web crawling methods (Mukhi et al., 2016). This source can provide early signals of disease emergence events to enhance management efforts by organizations such as GPHIN. The third big data source is Earth observation data obtained by satellites. Earth observation data provide proxies for environmental data such as weather, climate, habitats, and land use, which can be used in risk modelling to identify where there are risks of emergence and re-emergence of environmentally sensitive diseases (Ceccato et al., 2018). There are many examples of Earth observation data being used to identify disease risk (particularly Lyme disease) in Canada, in the form of static risk maps (Figure 6.9). Earth observation data are becoming increasingly “big” due to an increase in the numbers and types of satellites, with data at increasingly fine spatial resolution. In the near future, a higher frequency of images may allow for real-time identification of risks from vector-borne diseases such as WNV. At least theoretically, Earth observation data, synthesized by risk modelling to produce risk information could replace surveillance as a way to trigger public health responses well in advance of human cases of disease (Ogden et al., 2019).
Risk maps for Lyme disease in Eastern Canada based on the risk of Ixodes scapularis populations, using Earth observation data on climatic and habitat suitability.
Figure 6.9
These risk maps measure the environmental suitability for the tick vector of Lyme disease Ixodes scapularis in Eastern and Central Canada using Earth observation data proxies for temperature (annual cumulative degree-days above 0°C) and woodland habitat. The environmental suitability is zero where there is no woodland habitat (lighter grey areas) and where the climate is too cold (dark blue areas). In areas of woodland habitat, and where climate is warm enough, risk of occurrence of tick populations varies from low risk (pale blue areas) through moderate, moderate-high, and high-risk areas (represented by green, orange, and red areas, respectively). The scale of suitability for the tick according to temperature conditions (with values from 0 to 1500) is obtained from a mathematical model of I. scapularis populations. Earth observation data for this map were not available for parts of Nunavut and the Northwest Territories (shown by dark grey areas).
Source
Kotchi et al., 2019.
There has been much work done to identify and quantify the effects of weather on mosquito vectors of WNV and on WNV risk in Canada (see section 6.3.1.3 Canada-Endemic Mosquito-Borne Diseases), which would serve as a basis for developing forecasting of WNV outbreaks. Combining weather data and Earth observation data on habitat in predictive models in geographic information systems would allow forecasting at high spatiotemporal resolution.
Many of the challenges posed by emerging vectors and diseases that threaten public health in the context of climate change are shared by other disciplines, such as agriculture (including both plant and animal health), environment and biodiversity management, and natural resources, where species invasions (infectious or otherwise) will likely be driven by climate change, among other factors. Collaboration on risk assessment, surveillance, and management activities through a robust One Health approach may convey significant advantages over the siloed management of sector-specific risks (Ogden et al., 2019). This will require more systematic collaboration between disciplines and government departments, who are all dealing with the emergence of threats to health (human, animal, and environment) associated with climate change.
6.5
Knowledge Gaps
Share
Existing knowledge gaps related to the impact of climate change on infectious diseases in Canada hinder development of local and regional climate change and health vulnerability assessments and adaptation plans. They also thwart the ability of public health officials to detect infectious disease emergence and re-emergence, and to prevent and control such emergence. The key gaps are described below.
Greater knowledge is needed of the ecology and epidemiology of infectious diseases and arthropod vectors to support development of risk assessments and early warning systems. In addition, systematic surveillance of human cases, arthropod vectors, infected sentinel animals, and environmental samples, as well as use of citizen science methods, at useful spatiotemporal scales is needed to identify emerging diseases and vectors. While this is happening already to some extent, surveillance methods, such as metagenomics, that detect multiple, possibly emerging pathogens and vectors, are needed to move from surveillance targeting a single disease to programs that address broad ranges of pathogens and vectors that threaten Canadians. Infectious disease diagnostic methods and algorithms need to be developed so that diseases currently considered exotic to Canada are detected more readily in Canadians who have not travelled to countries where these diseases are historically known to occur. Surveillance should be long-term to support attribution of the effects of climate change.
There is a wide range of infectious diseases for which sensitivity to weather and climate is known, yet, for many of these, the possible impacts of climate change have not been assessed. In Canada, assessments that have been completed have focused on vector-borne diseases. The methods used for vector-borne diseases need to be applied to a wider range of infectious diseases, including human-to-human-transmitted diseases, and wind/airborne infectious diseases from environmental sources.
To attribute emergence and re-emergence of infectious diseases in Canada to climate change, information is required that supports analysis of statistical strength of association, consistency among studies, specificity of effects of climate, temporally appropriate timescales of climate change and disease emergence, biological gradient (greater effects of greater changes in climate), and plausibility (Hill, 1965).
There need to be greater linkages between the public health efforts to understand and respond to infectious and chronic diseases. It is increasingly recognized that infectious diseases can result in chronic illnesses (O’Connor et al., 2006), and that infectious diseases are more severe in those affected by chronic illnesses (Badawi et al., 2018). In addition, socio-economic status affects vulnerability to both infectious and chronic diseases as well as perceptions of risk, and knowledge of and willingness to use protective measures (Bouchard et al., 2018). The changing age demographic in Canada, as in other high-income countries, coupled with increasing incidence of chronic diseases in these populations (Dye, 2014), and the anticipated effects of climate change on both infectious and chronic diseases, mean that public health officials and researchers need to understand, consider, and respond to the impacts of climate change on these diseases in a coordinated fashion.
There are considerable gaps in Canada’s capacity to prevent and control vector-borne diseases. These gaps include a limited knowledge of a range of effective measures to control vectors in the environment and for people to protect themselves, as well as a lack of established protocols for vector-borne disease control programs for front-line public health responders. The need for innovation in prevention and control is highlighted by the increasing global challenges of antimicrobial resistance and resistance to insecticides.
Validation of early warning systems (Morin et al., 2018) is needed for those infectious diseases, such as mosquito-borne diseases, that may become more epidemic with climate change and that threaten Canada with rapid spread following introduction, as happened with WNV. In addition, Canadian communities at municipal and provincial or territorial levels require comprehensive assessments of vulnerability to the effects of climate change on all aspects of health, such as heat-related illnesses and deaths, chronic diseases, and risks from infectious diseases. These assessments are the first step to reducing future disease risks, as well as identifying knowledge and public health system gaps that need filling by investment and/or research (see Chapter 10: Health Sector Adaptation and Resilience).
Addressing existing knowledge gaps to support effective adaptation requires direct, well-planned, collaboration with public health and health end users (Nyström et al., 2018). Collaborations, such as that established between PHAC and Université de Montréal in their Groupe de recherche en épidémiologie des zoonoses et santé publique, would accelerate capacity to innovate and respond to emerging disease threats (GREZOSP, 2019). However, such collaborations require long-term investment; alignment of goals and objectives between partners; mutual skill, respect and trust; and geographical proximity to succeed (Rycroft-Malone et al., 2015).
6.6
Conclusion
Share
Since the first comprehensive Canadian assessment of climate change and health vulnerability and adaptation was published in 2008, there has been considerable effort to assess risks from emerging diseases (particularly vector-borne diseases), implement surveillance, and disseminate information on effective prevention and control to public health professionals and the public. Since that time, the tick vector of Lyme disease has spread into Canada, as projected, and carried with it the rapid emergence of Lyme and other tick-borne diseases. EEEV has also expanded its range into Canada, an outbreak of WNV occurred in Eastern Canada, and the mosquito species Ae. albopictus has become established in Southern Ontario. These events demonstrate increasing risks of northward spread of vectors and vector-borne diseases from the United States and of weather-driven epidemics of endemic mosquito-borne diseases, as identified in the 2008 assessment. They also highlight the reality of vector-borne disease emergence and re-emergence driven by climate change.
The emergence of, first, chikungunya virus infection and, then, Zika virus in the Americas has also increased concern among public health officials in and outside Canada of the risks associated with the spread of exotic diseases from far away and their introduction and endemic transmission. Whether these disease emergence events were driven by climate change, in whole or in part, has been the subject of debate (Paz & Semenza, 2016). Either way, the identification in Southern Ontario of populations of Ae. albopictus, a vector of both these diseases, means that the possibility of autochthonous transmission of exotic diseases in Canada is now more than speculation. Evidence presented in this chapter suggests that the most important exotic vector-borne diseases that could emerge in Canada by long-distance dispersal include dengue and Rift Valley fever. In addition, although plague and directly transmitted zoonoses such as hantavirus were identified as climate-sensitive diseases in the 2008 assessment, there have been no major emergence or re-emergence events associated with these diseases since that time. However, there have also been no efforts to further explore possible future effects of climate change on these pathogens.
Climate change and weather may increase risks from human-to-human-transmitted infectious diseases and airborne infectious diseases from environmental sources. Outbreaks or emergence events of these climate-sensitive diseases, such as acute flaccid paralysis and blastomycosis, have occurred in Canada or North America generally (Trudel et al., 2014; Elrick et al., 2019). Therefore, more research on the possible effects of climate change on these diseases is warranted.
Similar to other high-income countries, the adaptive capacity in Canada to manage emerging and re-emerging infectious disease risks is considered strong. This is due to the combined contribution of international and national public health systems, as well as robust health care services and technological capacity. Nevertheless, knowledge gaps and gaps in public health activities remain. These include the need for further risk assessments of the effects of climate change on infectious diseases; more systematic vulnerability assessments by municipalities, provinces, and territories; enhancements to surveillance, including long-term surveillance programs to monitor effects of climate change; and greater capacity for prevention and control. These gaps can be filled only by increased research and investment in public health responses, more direct collaboration between public health end users and academic researchers, and application of multidisciplinary One Health approaches.
Footnotes
Traditional Inuit food, also known as country food, is an integral part of Inuit identity and culture, a significant source of nutrients, and contributes to individual and community health and well-being. It includes marine animals (e.g. walrus, seals, etc.), caribou, birds, fish, and foraged foods.
↩
References
Acheson, E. S., Galanis, E., Bartlett, K., Klinkenberg, B. (2019). Climate classification system-based determination of temperate climate detection of Cryptococcus gattii sensu lato. Emerging Infectious Diseases, 25(9), 1723-1726.
↩
Acheson, E. S., Galanis, E., Bartlett, K., Mak, S., & Klinkenberg, B. (2018). Searching for clues for eighteen years: Deciphering the ecological determinants of Cryptococcus gattii on Vancouver Island, British Columbia. Medical Mycology, 56(2), 129-144. https://doi.org/10.1093/mmy/myx037 ↩
Adekoya N, Truman B, Landen M; Centers for Disease Control and Prevention. (2015). Incidence of notifiable diseases among American Indians/Alaska Natives – United States, 2007-2011. MMWR Morb Mortal Weekly Report, 64(1), 16-9.
↩
Adler NE, Newman K. (2002). Socioeconomic disparities in health: pathways and policies. Health Affairs (Project Hope), 21(2), 60-76. https://doi.org/10.1377/hlthaff.21.2.60 ↩
Aenishaenslin, C., Simon, A., Forde, T., Ravel, A., Proulx, J.-F., Fehlner-Gardiner, C., . . . Belanger, D. (2014). Characterizing rabies epidemiology in remote Inuit communities in Québec, Canada: A “One Health” approach. EcoHealth, 11(3), 343-355. https://doi.org/10.1007/s10393-014-0923-1 ↩
Alarcon Falconi, T. M., Cruz, M. S., & Naumova, E. N. (2018). The shift in seasonality of legionellosis in the USA. Epidemiology & Infection, 146(14), 1824-1833. https://doi.org/10.1017/S0950268818002182 ↩
Altizer, S., Ostfeld, R. S., Johnson, P. T. J., Kutz, S., & Harvell, C. D. (2013). Climate change and infectious diseases: From evidence to a predictive framework. Science, 341(6145), 514-519. https://doi.org/10.1126/science.1239401 ↩
Ampofo, K., Gesteland, P. H., Bender, J., Mills, M., Daly, J., Samore, M., . . . Srivastava, R. (2006). Epidemiology, complications, and cost of hospitalization in children with laboratory-confirmed influenza infection. Pediatrics, 118(6), 2409-2417. https://doi.org/10.1542/peds.2006-1475 ↩
Andersen, K. G., Rambaut, A., Lipkin, W. I., Holmes, E. C., & Garry, R. F. (2020). The proximal origin of SARS-CoV-2. Nature medicine, 26(4), 450–452. https://doi.org/10.1038/s41591-020-0820-9 ↩
Anderson, H., Honish, L., Taylor, G., Johnson, M., Tovstiuk, C., Fanning, A., Tyrrell, G., Rennie, R., Jaipaul, J., Sand, C., & Probert, S. (2006). Histoplasmosis cluster, golf course, Canada. Emerging infectious diseases, 12(1), 163–165. https://doi.org/10.3201/eid1201.051083 ↩
Anderson, R. M., & May, R. M. (1991). Infectious diseases of humans: Dynamics and control. Oxford, England: Oxford University Press.
↩
Andreadis, T. G., Anderson, J. F., Armstrong, P. M., & Main, A. J. (2008). Isolations of Jamestown Canyon virus (Bunyaviridae: Orthobunyavirus) from field-collected mosquitoes (Diptera: Culicidae) in Connecticut, USA: A ten-year analysis, 1997–2006. Vector-Borne and Zoonotic Diseases, 8(2), 175-188. https://doi.org/10.1089/vbz.2007.0169 ↩
Anyamba, A., Linthicum, K. J., Small, J. L., Collins, K. M., Tucker, C. J., Pak, E. W., . . . Russell, K. L. (2012). Climate teleconnections and recent patterns of human and animal disease outbreaks. PLOS Neglected Tropical Diseases, 6(1), 1-14. https://doi.org/10.1371/journal.pntd.0001465 ↩
Arksey, H., & O’Malley, L. (2005). Scoping studies: Towards a methodological framework. International Journal of Social Research Methodology, 8(1), 19-32. https://doi.org/10.1080/1364557032000119616 ↩
Armstrong, P. M., & Andreadis, T. G. (2010). Eastern equine encephalitis virus in mosquitoes and their role as bridge vectors. Emerging Infectious Diseases, 16(12), 1869-1874. https://doi.org/10.3201/eid1612.100640 ↩
Artsob, H. (1988). Powassan encephalitis. In T. P. Monath (Ed.), The arboviruses: Epidemiology and ecology (Vol. 4, pp. 29-49). Boca Raton, FL: CRC Press.
↩
Artsob, H. (2000). Arthropod-borne disease in Canada: A clinician’s perspective from the ‘Cold Zone’. Paediatrics & Child Health, 5(4), 206-212. https://doi.org/10.1093/pch/5.4.206 ↩
Artsob, H., Gubler, D. J., Enria, D. A., Morales, M. A., Pupo, M., Bunning, M. L., & Dudley, J. P. (2009). West Nile virus in the New World: Trends in the spread and proliferation of West Nile virus in the Western Hemisphere. Zoonoses and Public Health, 56(6-7), 357-369. https://doi.org/10.1111/j.1863-2378.2008.01207.x ↩
Asad, H., & Carpenter, D. O. (2018). Effects of climate change on the spread of Zika virus: A public health threat. Reviews on Environmental Health, 33(1), 31-42. https://doi.org/10.1515/reveh-2017-0042 ↩
Ashraf, U., Ye, J., Ruan, X., Wan, S., Zhu, B., & Cao, S. (2015). Usutu virus: An emerging flavivirus in Europe. Viruses, 7(1), 219-238. https://doi.org/10.3390/v7010219 ↩
Aune, K., Rhyan, J. C., Russell, R., Roffe, T. J., & Corso, B. (2012). Environmental persistence of Brucella abortus in the Greater Yellowstone Area. The Journal of Wildlife Management, 76(2), 253-261. https://doi.org/10.1002/jwmg.274 ↩
Ayers, V. B., Huang, Y.-J. S., Lyons, A. C., Park, S. L., Higgs, S., Dunlop, J. I., . . . Vanlandingham, D. L. (2018). Culex tarsalis is a competent vector species for Cache Valley virus. Parasites & Vectors, 11, 1-6. https://doi.org/10.1186/s13071-018-3103-2 ↩
Azam, D., Ukpai, O. M., Said, A., Abd-Allah, G. A., & Morgan, E. R. (2012). Temperature and the development and survival of infective Toxocara canis larvae. Parasitology Research, 110(2), 649-656. https://doi.org/10.1007/s00436-011-2536-8 ↩
Azar, M. M., Assi, R., Relich, R. F., Schmitt, B. H., Norris, S., Wheat, L. J., & Hage, C. A. (2015). Blastomycosis in Indiana. Chest, 148(5), 1276-1284. https://doi.org/10.1378/chest.15-0289 ↩
Badawi, A., Velummailum, R., Ryoo, S. G., Senthinathan, A., Yaghoubi, S., Vasileva, D., . . . Arora, P. (2018). Prevalence of chronic comorbidities in dengue fever and West Nile virus: A systematic review and meta-analysis. PLOS ONE, 13(7), 1-27. https://doi.org/10.1371/journal.pone.0200200 ↩
Baumgardner, D. J., & Burdick, J. S. (1991). An outbreak of human and canine blastomycosis. Reviews of Infectious Diseases, 13(5), 898-905. https://doi.org/10.1093/clinids/13.5.898 ↩
Baumgardner, D. J., Paretsky, D. P., Baeseman, Z. J., & Schreiber, A. (2011). Effects of season and weather on blastomycosis in dogs: Northern Wisconsin, USA. Medical Mycology, 49(1), 49-55. https://doi.org/10.3109/13693786.2010.488658 ↩
Beard, C. B., Eisen, R. J., Barker, C. M., Garofalo, J. F., Hahn, M., Hayden, M., . . . Schramm, P. J. (2016). Vector-borne diseases. In A. Crimmins, J. Balbus, J. L. Gamble, C. B. Beard, J. E. Bell, D. Dodgen, R. J. Eisen, N. Fann, M. D. Hawkins, S. C. Herring, L. Jantarasami, D. M. Mills, S. Saha, M. C. Sarofim, J. Trtanj, & L. Ziska (Eds.), The impacts of climate change on human health in the United States: A scientific assessment (pp. 129-156). Washington, DC: U.S. Global Change Research Program. https://doi.org/10.7930/J0765C7V ↩
Beaudoin, M., & Gosselin, P. (2016). An effective public health program to reduce urban heat islands in Québec, Canada. Pan American Journal of Public Health, 40(3), 160-166. Retrieved from http://iris.paho.org/xmlui/handle/123456789/31230 ↩
Beauté, J., Sandin, S., Uldum, S. A., Rota, M. C., Brandsema, P., Giesecke, J., & Sparén, P. (2016). Short-term effects of atmospheric pressure, temperature, and rainfall on notification rate of community-acquired Legionnaires’ disease in four European countries. Epidemiology & Infection, 144(16), 3483-3493. https://doi.org/10.1017/S0950268816001874 ↩
Ben Ari, T., Gershunov, A., Gage, K. L., Snäll, T., Ettestad, P., Kausrud, K. L., & Stenseth, N. C. (2008). Human plague in the USA: The importance of regional and local climate. Biology Letters, 4(6), 737-740. https://doi.org/10.1098/rsbl.2008.0363 ↩
Ben Ari, T., Gershunov, A., Tristan, R., Cazelles, B., Gage, K., & Stenseth, N. C. (2010). Interannual variability of human plague occurrence in the western United States explained by tropical and North Pacific Ocean climate variability. The American Journal of Tropical Medicine and Hygiene, 83(3), 624-632. https://doi.org/10.4269/ajtmh.2010.09-0775 ↩
Bern, C., & Montgomery, S. P. (2009). An estimate of the burden of Chagas disease in the United States. Clinical Infectious Diseases, 49(5), e52-e54. https://doi.org/10.1086/605091 ↩
Berrang-Ford, L., Maclean, J. D., Gyorkos, T. W., Ford, J. D., & Ogden, N. H. (2009). Climate change and malaria in Canada: A systems approach. Interdisciplinary Perspectives on Infectious Diseases, 2009, 1-13. https://doi.org/10.1155/2009/385487 ↩
Berry, P. (2008). Vulnerabilities, adaptation and adaptive capacity in Canada. In J. Séguin (Ed.), Human health in a changing climate: A Canadian assessment of vulnerabilities and adaptive capacity (pp. 367-448). Ottawa, ON: Government of Canada. Retrieved from http://publications.gc.ca/pub?id=9.691579&sl=0 ↩
Berry, P., Clarke, K.-L., Fleury, M. D., & Parker, S. (2014a). Human health. In F. J. Warren & D. S. Lemmen (Eds.), Canada in a changing climate: Sector perspectives on impacts and adaptation (pp. 191-232). Ottawa, ON: Government of Canada. Retrieved from http://publications.gc.ca/pub?id=9.699408&sl=0 ↩
Berry, P., Patterson, J., Buse, C. (2014b). Assessment of Vulnerability to the Health Impacts of Climate Change in Middlesex-London. Retrieved from https://www.healthunit.com/climate-change ↩
Berteaux, D., de Blois, S., Angers, J.-F., Bonin, J., Casajus, N., Darveau, M., . . . Vescovi, L. (2010). The CC-Bio project: Studying the effects of climate change on Quebec biodiversity. Diversity, 2(11), 1181-1204. https://doi.org/10.3390/d2111181 ↩
Berteaux, D., & Stenseth, N. C. (2006). Measuring, understanding and projecting the effects of large-scale climatic variability on mammals. Climate Research, 32(2), 95-97. Retrieved from https://www.int-res.com/articles/cr_oa/c032p095.pdf ↩
Biggs, H. M., Behravesh, C. B., Bradley, K. K., Dahlgren, F. S., Drexler, N. A., Dumler, J. S., . . . Traeger, M. S. (2016). Diagnosis and management of tickborne rickettsial diseases: Rocky Mountain spotted fever and other spotted fever group Rickettsioses, Ehrlichioses, and Anaplasmosis – United States. MMWR Recommendations and Reports, 65(2), 1-44. https://doi.org/10.15585/mmwr.rr6502a1 ↩
Blancher, P., Cadman, M. D., Pond, B. A., Couturier, A. R., Dunn, E. H., Francis, C. M., & Rempel, R. S. (2008). Changes in bird distributions between atlases. In M. D. Cadman, D. A. Sutherland, G. G. Beck, D. Lepage, & A. R. Couturier (Eds.), Atlas of the breeding birds of Ontario, 2001-2005 (pp. 32-48). Toronto, ON: Bird Studies Canada, Environment Canada, Ontario Field Ornithologists, Ontario Ministry of Natural Resources, & Ontario Nature.
↩
Bouchard, C., Aenishaenslin, C., Rees, E. E., Koffi, J. K., Pelcat, Y., Ripoche, M., . . . Leighton, P. A. (2018). Integrated social-behavioral and ecological risk maps to prioritize local public health responses to Lyme disease. Environmental Health Perspectives, 126(4). https://doi.org/10.1289/EHP1943 ↩
Bouchard, C., Dibernardo, A., Koffi, J., Wood, H., Leighton, P. A., & Lindsay, L. R. (2019). Increased risk of tick-borne diseases with climate and environmental changes. Canada Communicable Disease Report, 45(4), 83-89. https://doi.org/10.14745/ccdr.v45i04a02 ↩
Bouchard, C., Leonard, E., Koffi, J. K., Pelcat, Y., Peregrine, A., Chilton, N., . . . Ogden, N. H. (2015). The increasing risk of Lyme disease in Canada. The Canadian Veterinary Journal, 56(7), 693-699. Retrieved from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4466818/ ↩
Bouri, N., Sell, T. K., Franco, C., Adalja, A. A., Henderson, D. A., & Hynes, N. A. (2012). Return of epidemic dengue in the United States: Implications for the public health practitioner. Public Health Reports, 127(3), 259-266. https://doi.org/10.1177/003335491212700305 ↩
Brachman, P. S., & Abrutyn, E. (Eds.). (2009). Bacterial infections of humans: Epidemiology and control (4th ed.). New York, NY: Springer.
↩
Brady, O. J., Johansson, M. A., Guerra, C. A., Bhatt, S., Golding, N., Pigott, D. M., . . . Hay, S. I. (2013). Modelling adult Aedes aegypti and Aedes albopictus survival at different temperatures in laboratory and field settings. Parasites & Vectors, 6, 1-12. https://doi.org/10.1186/1756-3305-6-351 ↩
Brierley, L., Vonhof, M. J., Olival, K. J., Daszak, P., & Jones, K. E. (2016). Quantifying global drivers of zoonotic bat viruses: A process-based perspective. The American Naturalist, 187(2), E53-E64. https://doi.org/10.1086/684391 ↩
Brook, R. K., Kutz, S. J., Veitch, A. M., Popko, R. A., Elkin, B. T., & Guthrie, G. (2009). Fostering community-based wildlife health monitoring and research in the Canadian North. EcoHealth, 6(2), 266-278. https://doi.org/10.1007/s10393-009-0256-7 ↩
Brown, E. M., McTaggart, L. R., Dunn, D., Pszczolko, E., Tsui, K. G., Morris, S. K., . . . Richardson, S. E. (2018). Epidemiology and geographic distribution of blastomycosis, histoplasmosis, and coccidioidomycosis, Ontario, Canada, 1990–2015. Emerging Infectious Diseases, 24(7), 1257-1266. https://doi.org/10.3201/eid2407.172063 ↩
Brownstein, J. S., Holford, T. R., & Fish, D. (2005a). Effect of climate change on Lyme disease risk in North America. EcoHealth, 2(1), 38-46. https://doi.org/10.1007/s10393-004-0139-x ↩
Brownstein, J. S., Skelly, D. K., Holford, T. R., & Fish, D. (2005b). Forest fragmentation predicts local scale heterogeneity of Lyme disease risk. Oecologia, 146(3), 469-475. https://doi.org/10.1007/s00442-005-0251-9 ↩
Bullard, J. M. P., Ahsanuddin, A. N., Perry, A. M., Lindsay, L. R., Iranpour, M., Dibernardo, A., & Van Caeseele, P. G. (2014). The first case of locally acquired tick-borne Babesia microti infection in Canada. Canadian Journal of Infectious Diseases and Medical Microbiology, 25(6), e87-e89. https://doi.org/10.1155/2014/209521 ↩
Bush, E. and Lemmen, D.S. (Eds.). (2019). Canada’s Changing Climate Report. Ottawa, ON: Government of Canada. Retrieved from https://changingclimate.ca/CCCR2019/ ↩
Cable, J., Barber, I., Boag, B., Ellison, A. R., Morgan, E. R., Murray, K., . . . Booth, M. (2017). Global change, parasite transmission and disease control: Lessons from ecology. Philosophical Transactions of the Royal Society B: Biological Sciences, 372(1719), 1-17. https://doi.org/10.1098/rstb.2016.0088 ↩
Caini, S., Spreeuwenberg, P., Donker, G., Korevaar, J., & Paget, J. (2018). Climatic factors and long-term trends of influenza-like illness rates in the Netherlands, 1970–2016. Environmental Research, 167, 307-313. https://doi.org/10.1016/j.envres.2018.07.035 ↩
Campbell, L. P., Luther, C., Moo-Llanes, D., Ramsey, J. M., Danis-Lozano, R., & Peterson, A. T. (2015). Climate change influences on global distributions of dengue and chikungunya virus vectors. Philosophical Transactions of the Royal Society B: Biological Sciences, 370(1665), 1-9. https://doi.org/10.1098/rstb.2014.0135 ↩
Carlsen, B., & Glenton, C. (2016). The swine flu vaccine, public attitudes, and researcher interpretations: A systematic review of qualitative research. BMC Health Services Research, 16, 1-15. https://doi.org/10.1186/s12913-016-1466-7 ↩
Carmona-Castro, O., Moo-Llanes, D. A., & Ramsey, J. M. (2018). Impact of climate change on vector transmission of Trypanosoma cruzi (Chagas, 1909) in North America. Medical and Veterinary Entomology, 32(1), 84-101. https://doi.org/10.1111/mve.12269 ↩
Castillo, C. G., Kauffman, C. A., & Miceli, M. H. (2016). Blastomycosis. Infectious Disease Clinics of North America, 30(1), 247-264. https://doi.org/10.1016/j.idc.2015.10.002 ↩
Ceccato, P., Ramirez, B., Manyangadze, T., Gwakisa, P., & Thomson, M. C. (2018). Data and tools to integrate climate and environmental information into public health. Infectious Diseases of Poverty, 7, 1-11. https://doi.org/10.1186/s40249-018-0501-9 ↩
Centers for Disease Control and Prevention. (2018a). La Crosse encephalitis: Epidemiology & geographic distribution. Retrieved from https://www.cdc.gov/lac/tech/epi.html ↩
Centre for Food-borne, Environmental and Zoonotic Infectious Diseases. (2017). Synopsis: Lyme Disease in Canada – A Federal Framework. Canada communicable disease report, 43(10), 212-214. https://doi.org/10.14745/ccdr.v43i10a04 ↩
Chambers, D., Périé, C., Casajus, N., & de Blois, S. (2013). Challenges in modelling the abundance of 105 tree species in eastern North America using climate, edaphic, and topographic variables. Forest Ecology and Management, 291, 20-29. https://doi.org/10.1016/j.foreco.2012.10.046 ↩
Charron, D., Fleury, M., Lindsay, L. R., Ogden, N., & Schuster, C. J. (2008). The impacts of climate change on water-, food-, vector-,and rodent-borne diseases. In J. Séguin (Ed.), Human health in a changing climate: A Canadian assessment of vulnerabilities and adaptive capacity (pp. 171-210). Ottawa, ON: Government of Canada. Retrieved from http://publications.gc.ca/pub?id=9.691579&sl=0 ↩
Chattopadhyay, I., Kiciman, E., Elliott, J. W., Shaman, J. L., & Rzhetsky, A. (2018). Conjunction of factors triggering waves of seasonal influenza. eLife, 7, 1-44. https://doi.org/10.7554/eLife.30756 ↩
Chen, C. C., Jenkins, E., Epp, T., Waldner, C., Curry, P. S., & Soos, C. (2013). Climate change and West Nile virus in a highly endemic region of North America. International Journal of Environmental Research and Public Health, 10(7), 3052-3071. https://doi.org/10.3390/ijerph10073052 ↩
Chen, N.-T., Chen, M.-J., Guo, C.-Y., Chen, K.-T., & Su, H.-J. (2014a). Precipitation increases the occurrence of sporadic Legionnaires’ disease in Taiwan. PLOS ONE, 9(12), 1-11. https://doi.org/10.1371/journal.pone.0114337 ↩
Chen, S. C.-A., Meyer, W., & Sorrell, T. C. (2014b). Cryptococcus gattii infections. Clinical Microbiology Reviews, 27(4), 980-1024. https://doi.org/10.1128/CMR.00126-13 ↩
Cheng, J., Wu, J., Xu, Z., Zhu, R., Wang, X., Li, K., . . . Su, H. (2014). Associations between extreme precipitation and childhood hand, foot and mouth disease in urban and rural areas in Hefei, China. Science of the Total Environment, 497-498, 484-490. https://doi.org/10.1016/j.scitotenv.2014.08.006 ↩
Choi, K. M., Christakos, G., & Wilson, M. L. (2006). El Niño effects on influenza mortality risks in the state of California. Public Health, 120(6), 505-516. https://doi.org/10.1016/j.puhe.2006.01.011 ↩
Chretien, J.-P., Anyamba, A., Bedno, S. A., Breiman, R. F., Sang, R., Sergon, K., . . . Linthicum, K. J. (2007). Drought-associated chikungunya emergence along coastal East Africa. The American Journal of Tropical Medicine and Hygiene, 76(3), 405-407. https://doi.org/10.4269/ajtmh.2007.76.405 ↩
Cilloniz, C., Ewig, S., Gabarrus, A., Ferrer, M., Puig de la Bella Casa, J., Mensa, J., & Torres, A. (2017). Seasonality of pathogens causing community-acquired pneumonia. Respirology, 22(4), 778-785. https://doi.org/10.1111/resp.12978 ↩
Click Lambert, R., Kolivras, K. N., Resler, L. M., Brewster, C. C., & Paulson, S. L. (2008). The potential for emergence of Chagas disease in the United States. Geospatial Health, 2(2), 227-239. https://doi.org/10.4081/gh.2008.246 ↩
Clow, K. M., Leighton, P. A., Ogden, N. H., Lindsay, L. R., Michel, P., Pearl, D. L., & Jardine, C. M. (2017). Northward range expansion of Ixodes scapularis evident over a short timescale in Ontario, Canada. PLOS ONE, 12(12), 1-15. https://doi.org/10.1371/journal.pone.0189393 ↩
Collier, C. G. (1992). Weather conditions prior to major outbreaks of meningococcol meningitis in the United Kingdom. International Journal of Biometeorology, 36(1), 18-29. https://doi.org/10.1007/BF01208730 ↩
Comrie, A. C., & Glueck, M. F. (2007). Assessment of climate-coccidioidomycosis model: Model sensitivity for assessing climatologic effects on the risk of acquiring coccidioidomycosis. Annals of the New York Academy of Sciences, 1111(1), 83-95. https://doi.org/10.1196/annals.1406.024 ↩
Conza, L., Casati, S., Limoni, C., & Gaia, V. (2013). Meteorological factors and risk of community-acquired Legionnaires’ disease in Switzerland: An epidemiological study. BMJ Open, 3(3). https://doi.org/10.1136/bmjopen-2012-002428 ↩
Corrin, T., Greig, J., Harding, S., Young, I., Mascarenhas, M., & Waddell, L. A. (2018). Powassan virus, a scoping review of the global evidence. Zoonoses and Public Health, 65(6), 595-624. https://doi.org/10.1111/zph.12485 ↩
Cox, R., Sanchez, J., & Revie, C. W. (2013). Multi-criteria decision analysis tools for prioritising emerging or re-emerging infectious diseases associated with climate change in Canada. PLOS ONE, 8(8). https://doi.org/10.1371/journal.pone.0068338 ↩
Crampton, T. L., Light, R. B., Berg, G. M., Meyers, M. P., Schroeder, G. C., Hershfield, E. S., & Embil, J. M. (2002). Epidemiology and clinical spectrum of blastomycosis diagnosed at Manitoba hospitals. Clinical Infectious Diseases, 34(10), 1310-1316. https://doi.org/10.1086/340049 ↩
Cross, P. C., Edwards, W. H., Scurlock, B. M., Maichak, E. J., & Rogerson, J. D. (2007). Effects of management and climate on elk brucellosis in the Greater Yellowstone Ecosystem. Ecological Applications, 17(4), 957-964. https://doi.org/10.1890/06-1603 ↩
Dalcin, D., & Ahmed, S. Z. (2015). Blastomycosis in northwestern Ontario, 2004 to 2014. Canadian Journal of Infectious Diseases and Medical Microbiology, 26(5), 259-262. https://doi.org/10.1155/2015/468453 ↩
Davies, J. L., Epp, T., & Burgess, H. J. (2013). Prevalence and geographic distribution of canine and feline blastomycosis in the Canadian prairies. The Canadian Veterinary Journal, 54(8), 753-760. Retrieved from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3711163/ ↩
Davis, N. C. (1932). The effect of various temperatures in modifying the extrinsic incubation period of the yellow fever virus in Aëdes aegypti. American Journal of Epidemiology, 16(1), 163-176. https://doi.org/10.1093/oxfordjournals.aje.a117853 ↩
DeFelice, N. B., Schneider, Z. D., Little, E., Barker, C., Caillouet, K. A., Campbell, S. R., . . . Shaman, J. (2018). Use of temperature to improve West Nile virus forecasts. PLoS computational biology, 14(3). https://doi.org/10.1371/journal.pcbi.1006047 ↩
Delisle, E., Rousseau, C., Broche, B., Leparc-Goffart, I., L’Ambert, G., Cochet, A., . . . Golliot, F. (2015). Chikungunya outbreak in Montpellier, France, September to October 2014. Eurosurveillance, 20(17), 1-6. https://doi.org/10.2807/1560-7917.ES2015.20.17.21108 ↩
De Man, H., Mughini Gras, L., Schimmer, B., Friesema, I. H. M., De Roda Husman, A. M., & Van Pelt, W. (2016). Gastrointestinal, influenza-like illness and dermatological complaints following exposure to floodwater: A cross-sectional survey in the Netherlands. Epidemiology & Infection, 144(7), 1445-1454. https://doi.org/10.1017/S0950268815002654 ↩
Deplazes, P., Rinaldi, L., Alvarez Rojas, C. A., Torgerson, P. R., Harandi, M. F., Romig, T., . . . Jenkins, E. J. (2017). Global distribution of alveolar and cystic echinococcosis. Advances in Parasitology, 95, 315-493. https://doi.org/10.1016/bs.apar.2016.11.001 ↩
Dergousoff, S. J., Galloway, T. D., Lindsay, L. R., Curry, P. S., & Chilton, N. B. (2013). Range expansion of Dermacentor variabilis and Dermacentor andersoni (Acari: Ixodidae) near their northern distributional limits. Journal of Medical Entomology, 50(3), 510-520. https://doi.org/10.1603/me12193 ↩
DesGranges, J.-L., & Morneau, F. (2010). Potential sensitivity of Québec’s breeding birds to climate change. Avian Conservation and Ecology, 5(2). https://doi.org/10.5751/ace-00410-050205 ↩
Dibernardo, A., Cote, T., Ogden, N. H., & Lindsay, L. R. (2014). The prevalence of Borrelia miyamotoi infection, and co-infections with other Borrelia spp. in Ixodes scapularis ticks collected in Canada. Parasites & Vectors, 7, 1-8. https://doi.org/10.1186/1756-3305-7-183 ↩
Dicker, R. C., Coronado, F. t., Koo, D., & Gibson Parrish, R., II. (Eds.). (2012). Principles of epidemiology in public health practice: An introduction to applied epidemiology and biostatistics (3rd ed.). Atlanta, GA: Centers for Disease Control and Prevention.
↩
Dolcé, P., Bélanger, M.-J., Tumanowicz, K., Gauthier, C. P., Jutras, P., Massé, R., . . . Artsob, H. (2003). Coxiella burnetii seroprevalence of shepherds and their flocks in the lower Saint-Lawrence River region of Quebec, Canada. Canadian Journal of Infectious Diseases, 14(2), 97-102. https://doi.org/10.1155/2003/504796 ↩
Domenech de Cellès, M., Arduin, H., Lévy-Bruhl, D., Georges, S., Souty, C., Guillemot, D., . . . Opatowski, L. (2019). Unraveling the seasonal epidemiology of pneumococcus. Proceedings of the National Academy of Sciences of the United States of America, 116(5), 1802-1807. https://doi.org/10.1073/pnas.1812388116 ↩
Dowell, S. F. (2001). Seasonal variation in host susceptibility and cycles of certain infectious diseases. Emerging Infectious Diseases, 7(3), 369-374. https://doi.org/10.3201/eid0703.010301 ↩
Drebot, M. A., Artsob, H., & Werker, D. (2000). Hantavirus pulmonary syndrome in Canada, 1989-1999. Canada Communicable Disease Report, 26(8), 65-69. Retrieved from http://publications.gc.ca/pub?id=9.507730&sl=0 ↩
Dworkin, M. S., Schwan, T. G., Anderson, D. E., & Borchardt, S. M. (2008). Tick-borne relapsing fever. Infectious Disease Clinics of North America, 22(3), 449-468. https://doi.org/10.1016/j.idc.2008.03.006 ↩
Dye, C. (2014). After 2015: Infectious diseases in a new era of health and development. Philosophical Transactions of the Royal Society B: Biological Sciences, 369(1645), 1-9. https://doi.org/10.1098/rstb.2013.0426 ↩
Eads, D. A., & Hoogland, J. L. (2017). Precipitation, climate change, and parasitism of prairie dogs by fleas that transmit plague. Journal of Parasitology, 103(4), 309-319. https://doi.org/10.1645/16-195 ↩
Earn, D. J. D., He, D., Loeb, M. B., Fonseca, K., Lee, B. E., & Dushoff, J. (2012). Effects of school closure on incidence of pandemic influenza in Alberta, Canada. Annals of Internal Medicine, 156(3), 173-181. https://doi.org/10.7326/0003-4819-156-3-201202070-00005 ↩
Earn, D. J. D., Ma, J., Poinar, H., Dushoff, J., Bolker, B. M. (2020). Acceleration of plague outbreaks in the second pandemic Proceedings of the National Academies of Science of the U S A, 117(44), 27703-27711.
↩
Ebi, K.L., Balbus J.M., Luber G., Bole A., Crimmins A., Glass G., Saha S., Shimamoto M.M., Trtanj J., & White-Newsome J.L. (2018). Human Health. In Reidmiller, D.R., C.W. Avery, D.R. Easterling, K.E. Kunkel, K.L.M. Lewis, T.K. Maycock, and B.C. Stewart (Eds.), Impacts, Risks, and Adaptation in the United States: Fourth National Climate Assessment, Volume II (pp. 572–603). Washington, DC: U.S. Global Change Research Program. doi: 10.7930/NCA4.2018.CH14
↩
Ebi, K. L., Kovats, R. S., & Menne, B. (2006a). An approach for assessing human health vulnerability and public health interventions to adapt to climate change. Environmental Health Perspectives, 114(12), 1930-1934. https://doi.org/10.1289/ehp.8430 ↩
Ebi, K. L., Lewis, N. D., & Corvalan, C. (2006b). Climate variability and change and their potential health effects in small island states: Information for adaptation planning in the health sector. Environmental Health Perspectives, 114(12), 1957-1963. https://doi.org/10.1289/ehp.8429 ↩
Ebi, K. L., Ogden, N. H., Semenza, J. C., & Woodward, A. (2017). Detecting and attributing health burdens to climate change. Environmental Health Perspectives, 125(8), 1-8. https://doi.org/10.1289/EHP1509 ↩
Edginton, S., Guan, T. H., Evans, G., & Srivastava, S. (2018). Human granulocytic anaplasmosis acquired from a blacklegged tick in Ontario. CMAJ, 190(12), E363-E366. https://doi.org/10.1503/CMAJ.171243 ↩
Eisen, R. J., Eisen, L., Ogden, N. H., & Beard, C. B. (2016). Linkages of weather and climate with Ixodes scapularis and Ixodes pacificus (Acari: Ixodidae), enzootic transmission of Borrelia burgdorferi, and Lyme disease in North America. Journal of Medical Entomology, 53(2), 250-261. https://doi.org/10.1093/jme/tjv199 ↩
Eldin, C., Mahamat, A., Djossou, F., & Raoult, D. (2015). Rainfall and sloth births in May, Q fever in July, Cayenne, French Guiana. The American Journal of Tropical Medicine and Hygiene, 92(5), 979-981. https://doi.org/10.4269/ajtmh.14-0751 ↩
Elrick, M. J., Gordon-Lipkin, E., Crawford, T. O., Van Haren, K., Messacar, K., Thornton, N., . . . Duggal, P. (2019). Clinical subpopulations in a sample of North American children diagnosed with acute flaccid myelitis, 2012-2016. JAMA Pediatrics, 173(2), 134-139. https://doi.org/10.1001/jamapediatrics.2018.4890 ↩
Embil, J. A., Tanner, C. E., Pereira, L. H., Staudt, M., Morrison, E. G., & Gualazzi, D. A. (1988). Seroepidemiologic survey of Toxocara canis infection in urban and rural children. Public Health, 102(2), 129-133. https://doi.org/10.1016/S0033-3506(88)80039-8 ↩
Enscore, R. E., Biggerstaff, B. J., Brown, T. L., Fulgham, R. E., Reynolds, P. J., Engelthaler, D. M., . . . Gage, K. L. (2002). Modeling relationships between climate and the frequency of human plague cases in the southwestern United States, 1960-1997. The American Journal of Tropical Medicine and Hygiene, 66(2), 186-196. https://doi.org/10.4269/ajtmh.2002.66.186 ↩
Escobar, L. E., Romero-Alvarez, D., Leon, R., Lepe-Lopez, M. A., Craft, M. E., Borbor-Cordova, M. J., & Svenning, J.-C. (2016). Declining prevalence of disease vectors under climate change. Scientific Reports, 6, 1-8. https://doi.org/10.1038/srep39150 ↩
Fields, B. S., Benson, R. F., & Besser, R. E. (2002). Legionella and Legionnaires’ disease: 25 years of investigation. Clinical Microbiology Reviews, 15(3), 506-526. https://doi.org/10.1128/cmr.15.3.506-526.2002 ↩
Findlater, A., & Bogoch, I. I. (2018). Human mobility and the global spread of infectious diseases: A focus on air travel. Trends in Parasitology, 34(9), 772-783. https://doi.org/10.1016/j.pt.2018.07.004 ↩
Fisman, D. N., Lim, S., Wellenius, G. A., Johnson, C., Britz, P., Gaskins, M., . . . Newbern, C. (2005). It’s not the heat, it’s the humidity: Wet weather increases legionellosis risk in the greater Philadelphia metropolitan area. The Journal of Infectious Diseases, 192(12), 2066-2073. https://doi.org/10.1086/498248 ↩
Fisman, D. N., Tuite, A. R., & Brown, K. A. (2016). Impact of El Niño Southern Oscillation on infectious disease hospitalization risk in the United States. Proceedings of the National Academy of Sciences of the United States of America, 113(51), 14589-14594. https://doi.org/10.1073/pnas.1604980113 ↩
Fitzhenry, R., Weiss, D., Cimini, D., Balter, S., Boyd, C., Alleyne, L., . . . Varma, J. K. (2017). Legionnaires’ disease outbreaks and cooling towers, New York City, New York, USA. Emerging Infectious Diseases, 23(11), 1769-1776. https://doi.org/10.3201/eid2311.161584 ↩
Fonseca, K., Prince, G. D., Bratvold, J., Fox, J. D., Pybus, M., Preksaitis, J. K., & Tilley, P. (2005). West Nile virus infection and conjunctival exposure. Emerging Infectious Diseases, 11(10), 1648-1649. https://doi.org/10.3201/eid1110.040212 ↩
Food and Agriculture Organization, & World Health Organization. (2014). Multicriteria-based ranking for risk management of food-borne parasites: Report of a joint FAO/WHO expert meeting, 3-7 September 2012, FAO headquarters, Rome, Italy (Microbiological risk assessment series 23). Geneva, Switzerland: World Health Organization. Retrieved from https://www.who.int/foodsafety/publications/mra_23/en/ ↩
Forbes, L. B. (1991). Isolates of Brucella suis biovar 4 from animals and humans in Canada, 1982-1990. The Canadian Veterinary Journal, 32(11), 686-688. Retrieved from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1481085/ ↩
Franco-Paredes, C., Chastain, D., Taylor, P., Stocking, S., & Sellers, B. (2017). Boar hunting and brucellosis caused by Brucella suis. Travel Med Infect Dis, 16, 18-22. https://doi.org/10.1016/j.tmaid.2017.03.006 ↩
Friel, S. (2019). Climate change and the people’s health (N. Krieger, Ed.). Oxford University Press. doi:10.1093/oso/9780190492731.001.0001
↩
Gabriele-Rivet, V., Arsenault, J., Badcock, J., Cheng, A., Edsall, J., Goltz, J., . . . Ogden, N. H. (2015). Different ecological niches for ticks of public health significance in Canada. PLOS ONE, 10(7), 1-19. https://doi.org/10.1371/journal.pone.0131282 ↩
Gallant, D., Slough, B. G., Reid, D. G., & Berteaux, D. (2012). Arctic fox versus red fox in the warming Arctic: Four decades of den surveys in north Yukon. Polar Biology, 35(9), 1421-1431. https://doi.org/10.1007/s00300-012-1181-8 ↩
Garcia-Solache, M. A., & Casadevall, A. (2010). Global warming will bring new fungal diseases for mammals. mBio, 1(1), 1-3. https://doi.org/10.1128/mBio.00061-10 ↩
Garcia-Vidal, C., Labori, M., Viasus, D., Simonetti, A., Garcia-Somoza, D., Dorca, J., . . . Carratalà, J. (2013). Rainfall is a risk factor for sporadic cases of Legionella pneumophila pneumonia. PLOS ONE, 8(4), 1-5. https://doi.org/10.1371/journal.pone.0061036 ↩
Gargan, T. P., II., Clark, G. G., Dohm, D. J., Turell, M. J., & Bailey, C. L. (1988). Vector potential of selected North American mosquito species for Rift Valley fever virus. The American Journal of Tropical Medicine and Hygiene, 38(2), 440-446. https://doi.org/10.4269/ajtmh.1988.38.440 ↩
Garroway, C. J., Bowman, J., Cascaden, T. J., Holloway, G. L., Mahan, C. G., Malcolm, J. R., . . . Wilson, P. J. (2010). Climate change induced hybridization in flying squirrels. Global Change Biology, 16(1), 113-121. https://doi.org/10.1111/j.1365-2486.2009.01948.x ↩
Garroway, C. J., Bowman, J., Holloway, G. L., Malcolm, J. R., & Wilson, P. J. (2011). The genetic signature of rapid range expansion by flying squirrels in response to contemporary climate warming. Global Change Biology, 17(5), 1760-1769. https://doi.org/10.1111/j.1365-2486.2010.02384.x ↩
Gasmi, S., Bouchard, C., Ogden, N. H., Adam-Poupart, A., Pelcat, Y., Rees, E. E., . . . Thivierge, K. (2018). Evidence for increasing densities and geographic ranges of tick species of public health significance other than Ixodes scapularis in Québec, Canada. PLOS ONE, 13(8), 1-15. https://doi.org/10.1371/JOURNAL.PONE.0201924 ↩
Gasmi, S., Ogden, N. H., Lindsay, L. R., Burns, S., Fleming, S., Badcock, J., . . . Koffi, J. K. (2017). Surveillance for Lyme disease in Canada: 2009-2015. Canada Communicable Disease Report, 43(10), 194-199. https://doi.org/10.14745/ccdr.v43i10a01 ↩
Gilbert, M., Slingenbergh, J., & Xiao, X. (2008). Climate change and avian influenza. OIE Scientific and Technical Review, 27(2), 459-466. https://doi.org/10.20506/rst.27.2.1821 ↩
Gilmour, M. W., Graham, M., Reimer, A., & Van Domselaar, G. (2013). Public health genomics and the new molecular epidemiology of bacterial pathogens. Public Health Genomics, 16(1-2), 25-30. https://doi.org/10.1159/000342709 ↩
Giordano, B. V., Gasparotto, A., & Hunter, F. F. (2015). A checklist of the 67 mosquito species of Ontario, Canada. Journal of the American Mosquito Control Association, 31(1), 101-103. https://doi.org/10.2987/14-6456r.1 ↩
Giordano, B. V., Kaur, S., & Hunter, F. F. (2017). West Nile virus in Ontario, Canada: A twelve-year analysis of human case prevalence, mosquito surveillance, and climate data. PLOS ONE, 12(8), 1-15. https://doi.org/10.1371/journal.pone.0183568 ↩
Giordano, B. V., Turner, K. W., & Hunter, F. F. (2018). Geospatial analysis and seasonal distribution of West Nile virus vectors (Diptera: Culicidae) in Southern Ontario, Canada. International Journal of Environmental Research and Public Health, 15(4), 1-18. https://doi.org/10.3390/ijerph15040614 ↩
Githeko, A. K., Lindsay, S. W., Confalonieri, U. E., & Patz, J. A. (2000). Climate change and vector-borne diseases: A regional analysis. Bulletin of the World Health Organization, 78(9), 1136-1147. Retrieved from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2560843/ ↩
Gleason, J. A., Kratz, N. R., Greeley, R. D., & Fagliano, J. A. (2016). Under the weather: Legionellosis and meteorological factors. EcoHealth, 13(2), 293-302. https://doi.org/10.1007/s10393-016-1115-y ↩
Global Burden of Disease 2015 Mortality and Causes of Death Collaborators. (2016). Global, regional, and national life expectancy, all-cause mortality, and cause-specific mortality for 249 causes of death, 1980–2015: A systematic analysis for the Global Burden of Disease Study 2015. The Lancet, 388(10053), 1459-1544. https://doi.org/10.1016/S0140-6736(16)31012-1 ↩
Golnar, A. J., Kading, R. C., & Hamer, G. L. (2018). Quantifying the potential pathways and locations of Rift Valley fever virus entry into the United States. Transboundary and Emerging Diseases, 65(1), 85-95. https://doi.org/10.1111/tbed.12608 ↩
Golnar, A. J., Turell, M. J., LaBeaud, A. D., Kading, R. C., & Hamer, G. L. (2014). Predicting the mosquito species and vertebrate species involved in the theoretical transmission of Rift Valley fever virus in the United States. PLOS Neglected Tropical Diseases, 8(9), 1-11. https://doi.org/10.1371/journal.pntd.0003163 ↩
Goneau, L. W., Mehta, K., Wong, J., L’Huillier, A. G., & Gubbay, J. B. (2018). Zoonotic influenza and human health — Part 1: Virology and epidemiology of zoonotic influenzas. Current Infectious Disease Reports, 20(10), 1-12. https://doi.org/10.1007/s11908-018-0642-9 ↩
Gorris, M. E., Cat, L. A., Zender, C. S., Treseder, K. K., & Randerson, J. T. (2018). Coccidioidomycosis dynamics in relation to climate in the southwestern United States. GeoHealth, 2(1), 6-24. https://doi.org/10.1002/2017GH000095 ↩
Granados, D. P., & Castañeda, E. (2006). Influence of climatic conditions on the isolation of members of the Cryptococcus neoformans species complex from trees in Colombia from 1992-2004. FEMS Yeast Research, 6(4), 636-644. https://doi.org/10.1111/j.1567-1364.2006.00090.x ↩
Greer, A., Ng, V., & Fisman, D. (2008). Climate change and infectious diseases in North America: The road ahead. CMAJ, 178(6), 715-722. https://doi.org/10.1503/cmaj.081325 ↩
Greer, A. L., Drews, S. J., & Fisman, D. N. (2009). Why “winter” vomiting disease? Seasonality, hydrology, and norovirus epidemiology in Toronto, Canada. EcoHealth, 6(2), 192-199. https://doi.org/10.1007/s10393-009-0247-8 ↩
Griot, C., Gonzalez-Scarano, F., & Nathanson, N. (1993). Molecular determinants of the virulence and infectivity of California serogroup bunyaviruses. Annual Review of Microbiology, 47, 117-138. https://doi.org/10.1146/annurev.mi.47.100193.001001 ↩
Groupe de recherche en épidémiologie des zoonoses et santé publique. (2019). Retrieved from https://grezosp.com/ ↩
Gubler, D. J. (2007). The continuing spread of West Nile virus in the Western Hemisphere. Clinical Infectious Diseases, 45(8), 1039-1046. https://doi.org/10.1086/521911 ↩
Guerra, M., Walker, E. D., Jones, C., Paskewitz, S., Cortinas, M. R., Stancil, A., . . . Kitron, U. (2002). Predicting the risk of Lyme disease: Habitat suitability for Ixodes scapularis in the North Central United States. Emerging Infectious Diseases, 8(3), 289-297. Retrieved from https://wwwnc.cdc.gov/eid/article/8/3/01-0166_article ↩
Gupta, S., Parker, J., Smits, S., Underwood, J., & Dolwani, S. (2020). Persistent viral shedding of SARS-CoV-2 in faeces – a rapid review. Colorectal disease: the official journal of the Association of Coloproctology of Great Britain and Ireland, 22(6), 611–620. https://doi.org/10.1111/codi.15138 ↩
Hajizadeh M, Mitnitski A, Rockwood K. 2016. Socioeconomic gradient in health in Canada: Is the gap widening or narrowing? Health Policy, 120(9), 1040-50. doi: 10.1016/j.healthpol.2016.07.019.
↩
Halsby, K. D., Joseph, C. A., Lee, J. V., & Wilkinson, P. (2014). The relationship between meteorological variables and sporadic cases of Legionnaires’ disease in residents of England and Wales. Epidemiology & Infection, 142(11), 2352-2359. https://doi.org/10.1017/S0950268813003294 ↩
Harrigan, R. J., Thomassen, H. A., Buermann, W., & Smith, T. B. (2014). A continental risk assessment of West Nile virus under climate change. Global Change Biology, 20(8), 2417-2425. https://doi.org/10.1111/gcb.12534 ↩
Hassel, C., Mirand, A., Lukashev, A., Terletskaia-Ladwig, E., Farkas, A., Schuffenecker, I., . . . Bailly, J.-L. (2015). Transmission patterns of human enterovirus 71 to, from and among European countries, 2003 to 2013. Eurosurveillance, 20(34), 1-11. https://doi.org/10.2807/1560-7917.ES.2015.20.34.30005 ↩
Heesterbeek, H., Anderson, R. M., Andreasen, V., Bansal, S., De Angelis, D., Dye, C., . . . Isaac Newton Institute IDD Collaboration. (2015). Modeling infectious disease dynamics in the complex landscape of global health. Science, 347(6227), 1-10. https://doi.org/10.1126/science.aaa4339 ↩
Henao-Restrepo, A. M., Camacho, A., Longini, I. M., Watson, C. H., Edmunds, W. J., Egger, M., . . . Kieny, M.-P. (2017). Efficacy and effectiveness of an rVSV-vectored vaccine in preventing Ebola virus disease: Final results from the Guinea ring vaccination, open-label, cluster-randomised trial (Ebola Ça Suffit!). The Lancet, 389(10068), 505-518. https://doi.org/10.1016/S0140-6736(16)32621-6 ↩
Henri, D.A., Jean-Gagnon, F., & Grant Gilchrist, H. (2018). Using Inuit traditional ecological knowledge for detecting and monitoring avian cholera among Common Eiders in the eastern Canadian Arctic. Ecology and Society, 23(1), 22.
↩
Hersteinsson, P., & MacDonald, D. W. (1992). Interspecific competition and the geographical distribution of red and arctic foxes Vulpes vulpes and Alopex lagopus. Oikos, 64(3), 505-515. https://doi.org/10.2307/3545168 ↩
Hertig, E. (2019). Distribution of Anopheles vectors and potential malaria transmission stability in Europe and the Mediterranean area under future climate change. Parasites & Vectors, 12, 1-9. https://doi.org/10.1186/s13071-018-3278-6 ↩
Hicks, L. A., Rose, C. E., Fields, B. S., Drees, M. L., Engel, J. P., Jenkins, P. R., . . . Whitney, C. G. (2007). Increased rainfall is associated with increased risk for legionellosis. Epidemiology & Infection, 135(5), 811-817. https://doi.org/10.1017/S0950268806007552 ↩
Hierlihy, C., Waddell, L., Young, I., Greig, J., Corrin, T., & Mascarenhas, M. (2019). A systematic review of individual and community mitigation measures for prevention and control of chikungunya virus. PLOS ONE, 14(2), 1-27. https://doi.org/10.1371/journal.pone.0212054 ↩
Hill A.B. (1965). The environment and disease: association or causation? Proceedings of the Royal Society of Medicine, 58(5), 295–300.
↩
Hitch, A. T., & Leberg, P. L. (2007). Breeding distributions of North American bird species moving north as a result of climate change. Conservation Biology, 21(2), 534-539. https://doi.org/10.1111/j.1523-1739.2006.00609.x ↩
Hoegh-Guldberg, O., Jacob, D., Taylor, M., Bindi, M., Brown, S., Camilloni, I., . . . Zhou, G. (2019). Impacts of 1.5°C of global warming on natural and human systems. In V. Masson-Delmotte, P. Zhai, H.-O. Pörtner, D. Roberts, J. Skea, P. R. Shukla, A. Pirani, W. Moufouma-Okia, C. Péan, R. Pidcock, S. Connors, J. B. R. Matthews, Y. Chen, X. Zhou, M. I. Gomis, E. Lonnoy, T. Maycock, M. Tignor, & T. Waterfield (Eds.), Global warming of 1.5°C: An IPCC special report. In Press. Retrieved from https://www.ipcc.ch/sr15/chapter/chapter-3/ ↩
Holm-Hansen, C. C., Midgley, S. E., & Fischer, T. K. (2016). Global emergence of enterovirus D68: a systematic review. The Lancet Infectious Diseases, 16(5), e64-e75. https://doi.org/10.1016/S1473-3099(15)00543-5 ↩
Holt, A. C., Salkeld, D. J., Fritz, C. L., Tucker, J. R., & Gong, P. (2009). Spatial analysis of plague in California: Niche modeling predictions of the current distribution and potential response to climate change. International Journal of Health Geographics, 8, 1-14. https://doi.org/10.1186/1476-072x-8-38 ↩
Hongoh, V., Berrang-Ford, L., Scott, M. E., & Lindsay, L. R. (2012). Expanding geographical distribution of the mosquito, Culex pipiens, in Canada under climate change. Applied Geography, 33, 53-62. https://doi.org/10.1016/j.apgeog.2011.05.015 ↩
Huettmann, F., Magnuson, E. E., & Hueffer, K. (2017). Ecological niche modeling of rabies in the changing Arctic of Alaska. Acta Veterinaria Scandinavica, 59, 1-11. https://doi.org/10.1186/s13028-017-0285-0 ↩
Hunter, M., Donnelly, C., Smart, D., Smyth, B., Menzies, F., & Hedderwick, S. (2015). Brucellosis in people with occupational cattle exposure in Northern Ireland: Clinical features of 53 cases. Journal of Infection, 70(1), 101-103. https://doi.org/10.1016/j.jinf.2014.08.004 ↩
Institut national de santé publique du Québec. (2018a). Observatoire multipartite québécois sur les zoonoses et l’adaptation aux changements climatiques. Retrieved from https://www.inspq.qc.ca/zoonoses/observatoire ↩
Institut national de santé publique du Québec. (2018b). Priorisation des zoonoses au Québec dans un contexte d’adaptation aux changements climatiques à l’aide d’un outil d’aide à la décision multicritère. Retrieved from https://www.inspq.qc.ca/publications/2432 ↩
Intergovernmental Panel on Climate Change (IPCC). (2014). Summary for policymakers. In Field, C.B., V.R. Barros, D.J. Dokken, K.J. Mach, M.D. Mastrandrea, T.E. Bilir, M. Chatterjee, K.L. Ebi, Y.O. Estrada, R.C. Genova, B. Girma, E.S. Kissel, A.N. Levy, S. MacCracken, P.R. Mastrandrea, and L.L. White (Eds.), Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects.Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, United Kingdom and New York, NY, USA: Cambridge University Press.
↩
Iranpour, M., Hebert, P., Dibernardo, A., & Lindsay, R. (2017). Ochlerotatus ventrovittis Dyar (Diptera: Culicidae): A new record for the mosquito fauna of Canada. Paper presented at the Joint Meeting of the Entomological Societies of Canada and Manitoba: October 22-25, 2017, Winnipeg, MB. Retrieved from https://home.cc.umanitoba.ca/~fieldspg/pdf/Proceedings/ESMproceedings_V73.pdf ↩
Iranpour, M., Turell, M. J., & Lindsay, L. R. (2011). Potential for Canadian mosquitoes to transmit Rift Valley fever virus. Journal of the American Mosquito Control Association, 27(4), 363-369. https://doi.org/10.2987/11-6169.1 ↩
Jenkins, E. J., Castrodale, L. J., de Rosemond, S. J. C., Dixon, B. R., Elmore, S. A., Gesy, K. M., . . . Thompson, R. C. A. (2013). Tradition and transition: Parasitic zoonoses of people and animals in Alaska, northern Canada, and Greenland. Advances in Parasitology, 82, 33-204. https://doi.org/10.1016/B978-0-12-407706-5.00002-2 ↩
Jenkins, E. J., Schurer, J. M., & Gesy, K. M. (2011). Old problems on a new playing field: Helminth zoonoses transmitted among dogs, wildlife, and people in a changing northern climate. Veterinary Parasitology, 182(1), 54-69. https://doi.org/10.1016/j.vetpar.2011.07.015 ↩
Jetten, T. H., & Focks, D. A. (1997). Potential changes in the distribution of dengue transmission under climate warming. The American Journal of Tropical Medicine and Hygiene, 57(3), 285-297. https://doi.org/10.4269/ajtmh.1997.57.285 ↩
Ji, W., Huh, K., Kang, M., Hong, J., Bae, G. H., Lee, R., Na, Y., Choi, H., Gong, S. Y., Choi, Y. H., Ko, K. P., Im, J. S., & Jung, J. (2020). Effect of Underlying Comorbidities on the Infection and Severity of COVID-19 in Korea: a Nationwide Case-Control Study. Journal of Korean medical science, 35(25), e237. https://doi.org/10.3346/jkms.2020.35.e237 ↩
Jones, B., & O’Neill, B. C. (2016). Spatially explicit global population scenarios consistent with the Shared Socioeconomic Pathways. Environmental Research Letters, 11(8), 1-10. https://doi.org/10.1088/1748-9326/11/8/084003 ↩
Jones, K. E., Patel, N. G., Levy, M. A., Storeygard, A., Balk, D., Gittleman, J. L., & Daszak, P. (2008). Global trends in emerging infectious diseases. Nature, 451(7181), 990-993. https://doi.org/10.1038/nature06536 ↩
Jüni, P., Rothenbühler, M., Bobos, P., Thorpe, K. E., da Costa, B. R., Fisman, D. N., Slutsky, A. S., & Gesink, D. (2020). Impact of climate and public health interventions on the COVID-19 pandemic: a prospective cohort study. CMAJ : Canadian Medical Association journal = journal de l’Association medicale canadienne, 192(21), E566–E573. https://doi.org/10.1503/cmaj.200920 ↩
Kamal, M., Kenawy, M. A., Rady, M. H., Khaled, A. S., & Samy, A. M. (2019). Mapping the global potential distributions of two arboviral vectors Aedes aegypti and Ae. albopictus under changing climate. PLOS ONE, 13(12), 1-21. https://doi.org/10.1371/journal.pone.0210122 ↩
Kanzawa, M., Spindler, H., Anglemyer, A., & Rutherford, G. W. (2020). Will Coronavirus Disease 2019 Become Seasonal? The Journal of infectious diseases, 222(5), 719–721. https://doi.org/10.1093/infdis/jiaa345 ↩
Kauffman, C. A. (2007). Histoplasmosis: A clinical and laboratory update. Clinical Microbiology Reviews, 20(1), 115-132. https://doi.org/10.1128/CMR.00027-06 ↩
Kausrud, K. L., Begon, M., Ari, T. B., Viljugrein. H., Esper, J., Büntgen, U., Leirs, H., Junge, C., Yang, B., Yang, M., Xu, L., Stenseth, N. C. (2010) Modeling the epidemiological history of plague in Central Asia: palaeoclimatic forcing on a disease system over the past millennium. BMC Biology, 8, 112. doi: 10.1186/1741-7007-8-112.
↩
Kausrud, K. L., Mysterud, A., Steen, H., Vik, J. O., Østbye, E., Cazelles, B., . . . Stenseth, N. C. (2008). Linking climate change to lemming cycles. Nature, 456(7218), 93-97. https://doi.org/10.1038/nature07442 ↩
KC, S., & Lutz, W. (2017). The human core of the shared socioeconomic pathways: Population scenarios by age, sex and level of education for all countries to 2100. Global Environmental Change, 42, 181-192. https://doi.org/10.1016/j.gloenvcha.2014.06.004 ↩
Keegan, J. D., & Holland, C. V. (2010). Contamination of the hair of owned dogs with the eggs of Toxocara spp. Veterinary Parasitology, 173(1-2), 161-164. https://doi.org/10.1016/j.vetpar.2010.06.010 ↩
Kepron, M. W., Schoemperlen, C. B., Hershfield, E. S., Zylak, C. J., & Cherniack, R. M. (1972). North American blastomycosis in Central Canada: A review of 36 cases. CMAJ, 106(3), 243-246. Retrieved from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1940364/ ↩
Kidd, S. E., Hagen, F., Tscharke, R. L., Huynh, M., Bartlett, K. H., Fyfe, M., Macdougall, L., Boekhout, T., Kwon-Chung, K. J., & Meyer, W. (2004). A rare genotype of Cryptococcus gattii caused the cryptococcosis outbreak on Vancouver Island (British Columbia, Canada). Proceedings of the National Academy of Sciences of the United States of America, 101(49), 17258–17263. https://doi.org/10.1073/pnas.0402981101 ↩
Kidd, S. E., Bach, P. J., Hingston, A. O., Mak, S., Chow, Y., MacDougall, L., . . . Bartlett, K. H. (2007a). Cryptococcus gattii dispersal mechanisms, British Columbia, Canada. Emerging Infectious Diseases, 13(1), 51-57. https://doi.org/10.3201/eid1301.060823 ↩
Kidd, S. E., Chow, Y., Mak, S., Bach, P. J., Chen, H., Hingston, A. O., . . . Bartlett, K. H. (2007b). Characterization of environmental sources of the human and animal pathogen Cryptococcus gattii in British Columbia, Canada, and the Pacific Northwest of the United States. Applied and Environmental Microbiology, 73(5), 1433-1443. https://doi.org/10.1128/AEM.01330-06 ↩
Kilpatrick, A. M., Dobson, A. D. M., Levi, T., Salkeld, D. J., Swei, A., Ginsberg, H. S., . . . Diuk-Wasser, M. A. (2017). Lyme disease ecology in a changing world: Consensus, uncertainty and critical gaps for improving control. Philosophical Transactions of the Royal Society B: Biological Sciences, 372(1722). https://doi.org/10.1098/rstb.2016.0117 ↩
Kilpatrick, A. M., Kramer, L. D., Jones, M. J., Marra, P. P., & Daszak, P. (2006). West Nile virus epidemics in North America are driven by shifts in mosquito feeding behavior. PLOS Biology, 4(4), 0606-0610. https://doi.org/10.1371/journal.pbio.0040082 ↩
Kim, B. I., Ki, H., Park, S., Cho, E., & Chun, B. C. (2016). Effect of climatic factors on hand, foot, and mouth disease in South Korea, 2010-2013. PLOS ONE, 11(6), 1-9. https://doi.org/10.1371/journal.pone.0157500 ↩
Kinlin, L. M., Spain, C. V., Ng, V., Johnson, C. C., White, A. N. J., & Fisman, D. N. (2009). Environmental exposures and invasive meningococcal disease: An evaluation of effects on varying time scales. American Journal of Epidemiology, 169(5), 588-595. https://doi.org/10.1093/aje/kwn383 ↩
Kissler, S. M., Tedijanto, C., Goldstein, E., Grad, Y. H., & Lipsitch, M. (2020). Projecting the transmission dynamics of SARS-CoV-2 through the postpandemic period. Science (New York, N.Y.), 368(6493), 860–868. https://doi.org/10.1126/science.abb5793 ↩
Knobel, D. L., Cleaveland, S., Coleman, P. G., Fèvre, E. M., Meltzer, M. I., Miranda, M. E. G., . . . Meslin, F.-X. (2005). Re-evaluating the burden of rabies in Africa and Asia. Bulletin of the World Health Organization, 83(5), 360-368. Retrieved from https://www.who.int/bulletin/volumes/83/5/360.pdf ↩
Kotchi S.-O., Bouchard, C., Brazeau, S., Ogden, N.H. (2021) Earth Observation-informed risk maps of the Lyme disease vector Ixodes scapularis in Central and Eastern Canada. Remote Sensing, 13(3), 524.
↩
Kotwa, J. D., Isaksson, M., Jardine, C. M., Campbell, G. D., Berke, O., Pearl, D. L., . . . Peregrine, A. S. (2019). Echinococcus multilocularis infection, Southern Ontario, Canada. Emerging Infectious Diseases, 25(2), 265-272. https://doi.org/10.3201/eid2502.180299 ↩
Kraemer, M. U. G., Reiner, R. C., Jr., Brady, O. J., Messina, J. P., Gilbert, M., Pigott, D. M., . . . Golding, N. (2019). Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopictus. Nature Microbiology, 4, 854-863. https://doi.org/10.1038/s41564-019-0376-y ↩
Kraemer, M. U. G., Sinka, M. E., Duda, K. A., Mylne, A., Shearer, F. M., Brady, O. J., . . . Hay, S. I. (2015). The global compendium of Aedes aegypti and Ae. albopictus occurrence. Scientific Data, 2, 1-8. https://doi.org/10.1038/sdata.2015.35 ↩
Kulkarni, M. A., Berrang-Ford, L., Buck, P. A., Drebot, M. A., Lindsay, L. R., & Ogden, N. H. (2015). Major emerging vector-borne zoonotic diseases of public health importance in Canada. Emerging Microbes & Infections, 4(1), 1-7. https://doi.org/10.1038/emi.2015.33 ↩
Kuster, S. P., Tuite, A. R., Kwong, J. C., McGeer, A., The Toronto Invasive Bacterial Diseases Network, & Fisman, D. N. (2011). Evaluation of coseasonality of influenza and invasive pneumococcal disease: Results from prospective surveillance. PLOS Medicine, 8(6), 1-9. https://doi.org/10.1371/journal.pmed.1001042 ↩
Kwon-Chung, K. J., & Bennett, J. E. (1984). High prevalence of Cryptococcus neoformans var. gattii in tropical and subtropical regions. Zentralblatt für Bakteriologie, Mikrobiologie, und Hygiene Serie A – Medical microbiology, infectious diseases, virology, parasitology, 257(2), 213-218.
↩
Larrieu, S., Cassadou, S., Rosine, J., Chappert, J. L., Blateau, A., Ledrans, M., & Quénel, P. (2014). Lessons raised by the major 2010 dengue epidemics in the French West Indies. Acta Tropica, 131, 37-40. https://doi.org/10.1016/j.actatropica.2013.11.023 ↩
Larsen, J. N., Anisimov, O. A., Constable, A., Hollowed, A. B., Maynard, N., Prestrud, P., . . . Stone, J. M. R. (2014). Polar regions. In V. R. Barros, C. B. Field, D. J. Dokken, M. D. Mastrandrea, K. J. Mach, T. E. Bilir, M. Chatterjee, K. L. Ebi, Y. O. Estrada, R. C. Genova, B. Girma, E. S. Kissel, A. N. Levy, S. MacCracken, P. R. Mastrandrea, & L. L. White (Eds.), Climate change 2014: Impacts, adaptation, and vulnerability. Part B: Regional aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (pp. 1567-1612). Cambridge, United Kingdom: Cambridge University Press. Retrieved from https://www.ipcc.ch/report/ar5/wg2/ ↩
Lau, L., Wudel, B., Kadkhoda, K., & Keynan, Y. (2017). Snowshoe hare virus causing meningoencephalitis in a young adult from northern Manitoba, Canada. Open Forum Infectious Diseases, 4(3), 1-3. https://doi.org/10.1093/ofid/ofx150 ↩
Lau, S., Luk, H., Wong, A., Li, K., Zhu, L., He, Z., Fung, J., Chan, T., Fung, K., & Woo, P. (2020). Possible Bat Origin of Severe Acute Respiratory Syndrome Coronavirus 2. Emerging infectious diseases, 26(7), 1542–1547. https://doi.org/10.3201/eid2607.200092 ↩
Lee, A. C. Y., Schantz, P. M., Kazacos, K. R., Montgomery, S. P., & Bowman, D. D. (2010). Epidemiologic and zoonotic aspects of ascarid infections in dogs and cats. Trends in Parasitology, 26(4), 155-161. https://doi.org/10.1016/j.pt.2010.01.002 ↩
Leighton, P. A., Koffi, J. K., Pelcat, Y., Lindsay, L. R., & Ogden, N. H. (2012). Predicting the speed of tick invasion: An empirical model of range expansion for the Lyme disease vector Ixodes scapularis in Canada. Journal of Applied Ecology, 49(2), 457-464. https://doi.org/10.1111/j.1365-2664.2012.02112.x ↩
Levac, D., Colquhoun, H., & O’Brien, K. K. (2010). Scoping studies: Advancing the methodology. Implementation Science, 5, 69. https://doi.org/10.1186/1748-5908-5-69 ↩
Levine RS, Hedeen DL, Hedeen MW, Hamer GL, Mead DG, Kitron UD. (2017). Avian species diversity and transmission of West Nile virus in Atlanta, Georgia. Parasites & Vectors, 10(1), 62. doi: 10.1186/s13071-017-1999-6.
↩
Liao, J., Yu, S., Yang, F., Yang, M., Hu, Y., & Zhang, J. (2016). Short-term effects of climatic variables on hand, foot, and mouth disease in mainland China, 2008-2013: A multilevel spatial Poisson regression model accounting for overdispersion. PLOS ONE, 11(1), 1-14. https://doi.org/10.1371/journal.pone.0147054 ↩
Light, R. B., Kralt, D., Embil, J. M., Trepman, E., Wiebe, L., Limerick, B., . . . Macdonald, K. (2008). Seasonal variations in the clinical presentation of pulmonary and extrapulmonary blastomycosis. Medical Mycology, 46(8), 835-841. https://doi.org/10.1080/13693780802132763 ↩
Likos, A., Griffin, I., Bingham, A. M., Stanek, D., Fischer, M., White, S., . . . Philip, C. (2016). Local mosquito-borne transmission of Zika virus—Miami-Dade and Broward counties, Florida, June–August 2016. Morbidity and Mortality Weekly Report, 65(38), 1032-1038. https://doi.org/10.15585/mmwr.mm6538e1 ↩
Lin, H., Zou, H., Wang, Q., Liu, C., Lang, L., Hou, X., & Li, Z. (2013). Short-term effect of El Niño-Southern Oscillation on pediatric hand, foot and mouth disease in Shenzhen, China. PLOS ONE, 8(7), 1-7. http://doi.org/10.1371/journal.pone.0065585 ↩
Lindquist, E. E., Galloway, T. D., Artsob, H., Lindsay, L. R., Drebot, M., Wood, H., & Robbins, R. G. (2016). A handbook to the ticks of Canada (Ixodida: Ixodidae, Argasidae): Biological Survey of Canada Monograph Series No. 7: Biological Survey of Canada. Retrieved from https://biologicalsurvey.ca/public/Bsc/Controller/Page/AGR-001-Ticks-Monograph.pdf ↩
Lindsay, A. P., Hope, V., Marshall, R. J., & Salinger, J. (2002). Meningococcal disease and meteorological conditions in Auckland, New Zealand. Australian and New Zealand Journal of Public Health, 26(3), 212-218. https://doi.org/10.1111/j.1467-842X.2002.tb00676.x ↩
Lindsay, L. R., Barker, I. K., Surgeoner, G. A., Mcewen, S. A., Gillespie, T. J., & Addison, E. M. (1998). Survival and development of the different life stages of Ixodes scapularis (Acari: Ixodidae) held within four habitats on Long Point, Ontario, Canada. Journal of Medical Entomology, 35(3), 189-199. https://doi.org/10.1093/jmedent/35.3.189 ↩
Lindsey, N. P., Staples, J. E., & Fischer, M. (2018). Chikungunya virus disease among travelers—United States, 2014-2016. The American Journal of Tropical Medicine and Hygiene, 98(1), 192-197. https://doi.org/10.4269/ajtmh.17-0668 ↩
Litvinjenko, S., & Lunny, D. (2017). Blastomycosis hospitalizations in northwestern Ontario: 2006-2015. Canada Communicable Disease Report, 43(10), 200-205. https://doi.org/10.14745/ccdr.v43i10a02 ↩
Litvinov, I. V., St-Germain, G., Pelletier, R., Paradis, M., & Sheppard, D. C. (2013). Endemic human blastomycosis in Quebec, Canada, 1988–2011. Epidemiology & Infection, 141(6), 1143-1147. https://doi.org/10.1017/S0950268812001860 ↩
Litvintseva, A. P., Marsden-Haug, N., Hurst, S., Hill, H., Gade, L., Driebe, E. M., . . . Chiller, T. (2015). Valley fever: Finding new places for an old disease: Coccidioides immitis found in Washington State soil associated with recent human infection. Clinical Infectious Diseases, 60(1), e1-e3. https://doi.org/10.1093/cid/ciu681 ↩
Lohrenz, S., Minion, J., Pandey, M., & Karunakaran, K. (2018). Blastomycosis in southern Saskatchewan 2000–2015: Unique presentations and disease characteristics. Medical Mycology, 56(7), 787-795. https://doi.org/10.1093/mmy/myx131 ↩
Lopman, B., Armstrong, B., Atchison, C., & Gray, J. J. (2009). Host, weather and virological factors drive norovirus epidemiology: Time-series analysis of laboratory surveillance data in England and Wales. PLOS ONE, 4(8), 1-10. https://doi.org/10.1371/journal.pone.0006671 ↩
Ludwig, A., Bigras-Poulin, M., Michel, P., & Bélanger, D. (2010). Risk factors associated with West Nile virus mortality in American Crow populations in southern Quebec. Journal of Wildlife Diseases, 46(1), 195-208. https://doi.org/10.7589/0090-3558-46.1.195 ↩
Ludwig, A., Zheng, H., Vrbova, L., Drebot, M. A., Iranpour, M., & Lindsay, L. R. (2019). Increased risk of endemic mosquito-borne diseases in Canada due to climate change. Canada Communicable Disease Report, 45(4), 90-97. https://doi.org/10.14745/ccdr.v45i04a03 ↩
MacDougall, L., Fyfe, M., Romney, M., Starr, M., & Galanis, E. (2011). Risk factors for Cryptococcus gattii infection, British Columbia, Canada. Emerging Infectious Diseases, 17(2), 193-199. https://doi.org/10.3201/eid1702.101020 ↩
MacDougall, L., Kidd, S. E., Galanis, E., Mak, S., Leslie, M. J., Cieslak, P. R., . . . Bartlett, K. H. (2007). Spread of Cryptococcus gattii in British Columbia, Canada, and detection in the Pacific Northwest, USA. Emerging Infectious Diseases, 13(1), 42-50. https://doi.org/10.3201/eid1301.060827 ↩
Machado-Machado, E. A. (2012). Empirical mapping of suitability to dengue fever in Mexico using species distribution modeling. Applied Geography, 33, 82-93. https://doi.org/10.1016/j.apgeog.2011.06.011 ↩
Maiga, A. W., Deppen, S., Scaffidi, B. K., Baddley, J., Aldrich, M. C., Dittus, R. S., & Grogan, E. L. (2018). Mapping Histoplasma capsulatum exposure, United States. Emerging Infectious Diseases, 24(10), 1835-1839. https://doi.org/10.3201/eid2410.180032 ↩
Mak, S., Morshed, M., & Henry, B. (2010). Ecological niche modeling of Lyme disease in British Columbia, Canada. Journal of Medical Entomology, 47(1), 99-105. https://doi.org/10.1603/033.047.0114 ↩
Maksimovic, Z., Cornwell, M. S., Semren, O., & Rifatbegovic, M. (2017). The apparent role of climate change in a recent anthrax outbreak in cattle. OIE Scientific and Technical Review, 36(3), 959-963. https://doi.org/10.20506/rst.36.3.2727 ↩
Marsden-Haug, N., Goldoft, M., Ralston, C., Limaye, A. P., Chua, J., Hill, H., . . . Chiller, T. (2013). Coccidioidomycosis acquired in Washington state. Clinical Infectious Diseases, 56(6), 847-850. https://doi.org/10.1093/cid/cis1028 ↩
Marston, B. J., Lipman, H. B., & Breiman, R. F. (1994). Surveillance for Legionnaires’ disease: Risk factors for morbidity and mortality. Archives of Internal Medicine, 154(21), 2417-2422. https://doi.org/10.1001/archinte.1994.00420210049006 ↩
Mas-Coma, S., Valero, M. A., & Bargues, M. D. (2008). Effects of climate change on animal and zoonotic helminthiasis. OIE Scientific and Technical Review, 27(2), 443-457. https://doi.org/10.20506/rst.27.2.1822 ↩
McBride, J. A., Gauthier, G. M., & Klein, B. S. (2017). Clinical manifestations and treatment of blastomycosis. Clinics in Chest Medicine, 38(3), 435-449. https://doi.org/10.1016/j.ccm.2017.04.006 ↩
McDonough, E. S., Wisniewski, T. R., Penn, L. A., Chan, D. M., & McNamara, W. J. (1976). Preliminary studies on conidial liberation of Blastomyces dermatitidis and Histoplasma capsulatum. Sabouraudia, 14(2), 199-204. https://doi.org/10.1080/00362177685190271 ↩
McMichael, C., Barnett, J., & McMichael, A. J. (2012). An ill wind? Climate change, migration, and health. Environmental Health Perspectives, 120(5), 646-654. https://doi.org/10.1289/ehp.1104375 ↩
McTaggart, L. R., Brown, E. M., & Richardson, S. E. (2016). Phylogeographic analysis of Blastomyces dermatitidis and Blastomyces gilchristii reveals an association with North American freshwater drainage basins. PLOS ONE, 11(7), 1-22. https://doi.org/10.1371/journal.pone.0159396 ↩
Medlock, J. M., & Vaux, A. G. C. (2011). Assessing the possible implications of wetland expansion and management on mosquitoes in Britain. European Mosquito Bulletin, 29, 38-65. Retrieved from http://e-m-b.org/sites/e-m-b.org/files/EMB%2829%2938-65.pdf ↩
Melles, S. J., Fortin, M. J., Lindsay, K., & Badzinski, D. (2011). Expanding northward: Influence of climate change, forest connectivity, and population processes on a threatened species’ range shift. Global Change Biology, 17(1), 17-31. https://doi.org/10.1111/j.1365-2486.2010.02214.x ↩
Michele, M., Alberto, M., Liana, S., & Francesco, D. (2006). Do environmental factors influence the occurrence of acute meningitis in industrialized countries? An epidemic of varying aetiology in Northern Italy. European Journal of Epidemiology, 21(6), 465-468. https://doi.org/10.1007/s10654-006-9013-1 ↩
Millins, C., Gilbert, L., Medlock, J., Hansford, K., Thompson, D. B. A., & Biek, R. (2017). Effects of conservation management of landscapes and vertebrate communities on Lyme borreliosis risk in the United Kingdom. Philosophical Transactions of the Royal Society B: Biological Sciences, 372(1722), 1-12. https://doi.org/10.1098/rstb.2016.0123 ↩
Minigan, J. N., Hager, H. A., Peregrine, A. S., & Newman, J. A. (2018). Current and potential future distribution of the American dog tick (Dermacentor variabilis, Say) in North America. Ticks and Tick-borne Diseases, 9(2), 354-362. https://doi.org/10.1016/j.ttbdis.2017.11.012 ↩
Mitchell, R., & Kandola, K. (2005). Rabies in the Northwest Territories Part 2: Rabies surveillance in the Northwest Territories. EpiNorth, 17, 4-6.
↩
Monaghan, A. J., Moore, S. M., Sampson, K. M., Beard, C. B., & Eisen, R. J. (2015). Climate change influences on the annual onset of Lyme disease in the United States. Ticks and Tick-borne Diseases, 6(5), 615-622. https://doi.org/10.1016/j.ttbdis.2015.05.005 ↩
Moreno-Madriñán, M. J., & Turell, M. (2018). History of mosquitoborne diseases in the United States and implications for new pathogens. Emerging Infectious Diseases, 24(5), 821-826. https://doi.org/10.3201/eid2405.171609 ↩
Morin, C. W., Semenza, J. C., Trtanj, J. M., Glass, G. E., Boyer, C., & Ebi, K. L. (2018). Unexplored opportunities: Use of climate- and weather-driven early warning systems to reduce the burden of infectious diseases. Current Environmental Health Reports, 5(4), 430-438. https://doi.org/10.1007/s40572-018-0221-0 ↩
Morral-Puigmal, C., Martínez-Solanas, È., Villanueva, C. M., & Basagaña, X. (2018). Weather and gastrointestinal disease in Spain: A retrospective time series regression study. Environment International, 121(1), 649-657. https://doi.org/10.1016/j.envint.2018.10.003 ↩
Morris, S. K., Brophy, J., Richardson, S. E., Summerbell, R., Parkin, P. C., Jamieson, F., . . . Ford-Jones, E. L. (2006). Blastomycosis in Ontario, 1994-2003. Emerging Infectious Diseases, 12(2), 274-279. https://doi.org/10.3201/eid1202.050849 ↩
Mouchtouri, V. A., Verykouki, E., Zamfir, D., Hadjipetris, C., Lewis, H. C., Hadjichristodoulou, C., & The EU SHIPSAN ACT Partnership. (2017). Gastroenteritis outbreaks on cruise ships: Contributing factors and thresholds for early outbreak detection. Eurosurveillance, 22(45), 1-8. https://doi.org/10.2807/1560-7917.ES.2017.22.45.16-00576 ↩
Muchaal, P. K., Parker, S., Meganath, K., Landry, L., & Aramini, J. (2015). Evaluation of a national pharmacy-based syndromic surveillance system. Canada Communicable Disease Report, 41(9), 203-208. https://doi.org/10.14745/ccdr.v41i09a01 ↩
Mukhi, S. N., Andres, E., Demianyk, B., Gammon, B., & Kloeze, H. (2016). KIWI: A technology for public health event monitoring and early warning detection. Online Journal of Public Health Informatics, 8(3), 1-16. https://doi.org/10.5210/ojphi.v8i3.6937 ↩
Murray, D. L. (2003). Relapsing fever. In C. D. Rudolph, A. M. Rudolph, M. K. Hostetter, G. Lister, & N. J. Siegel (Eds.), Rudolph’s pediatrics (21st ed.). New York, NY: McGraw-Hill.
↩
Nakazawa, Y., Williams, R., Peterson, A. T., Mead, P., Staples, E., & Gage, K. L. (2007). Climate change effects on plague and tularemia in the United States. Vector-Borne and Zoonotic Diseases, 7(4), 529-540. https://doi.org/10.1089/vbz.2007.0125 ↩
Nash, D., & Geng, E. (2020). Goal-Aligned, Epidemic Intelligence for the Public Health Response to the COVID-19 Pandemic. American journal of public health, 110(8), 1154–1156. https://doi.org/10.2105/AJPH.2020.305794 ↩
National Collaborating Centre for Aboriginal Health. (2016). Determinants of the prevalence and severity of influenza infection in Indigenous populations in Canada. Prince George, BC: National Collaborating Centre for Aboriginal Health.
↩
Naughton, D. (2012). The natural history of Canadian mammals. Toronto, ON: University of Toronto Press.
↩
Neil, K., & Berkelman, R. (2008). Increasing incidence of Legionellosis in the United States, 1990–2005: Changing epidemiologic trends. Clinical Infectious Diseases, 47(5), 591-599. https://doi.org/10.1086/590557 ↩
Nelder, M. P., Russell, C. B., Clow, K. M., Johnson, S., Weese, J. S., Cronin, K., . . . Patel, S. N. (2019). Occurrence and distribution of Ambylomma americanum as determined by passive surveillance in Ontario, Canada (1999–2016). Ticks and Tick-borne Diseases, 10(1), 146-155. https://doi.org/10.1016/J.TTBDIS.2018.10.001 ↩
Neumann, G. (2015). H5N1 influenza virulence, pathogenicity and transmissibility: What do we know? Future Virology, 10(8), 971-980. https://doi.org/10.2217/fvl.15.62 ↩
Neumann, G., Noda, T., & Kawaoka, Y. (2009). Emergence and pandemic potential of swine-origin H1N1 influenza virus. Nature, 459(7249), 931-939. https://doi.org/10.1038/nature08157 ↩
Ng, V., Fazil, A., Gachon, P., Deuymes, G., Radojević, M., Mascarenhas, M., . . . Ogden, N. H. (2017). Assessment of the probability of autochthonous transmission of chikungunya virus in Canada under recent and projected climate change. Environmental Health Perspectives, 125(6), 1-12. https://doi.org/10.1289/ehp669 ↩
Ng, V., Rees, E. E., Lindsay, R. L., Drebot, M. A., Brownstone, T., Sadeghieh, T., & Khan, S. U. (2019). Could exotic mosquito-borne diseases emerge in Canada with climate change? Canada Communicable Disease Report, 45(4), 98-107. https://doi.org/10.14745/ccdr.v45i04a04 ↩
Ng, V., & Sargeant, J. M. (2012). A stakeholder-informed approach to the identification of criteria for the prioritization of zoonoses in Canada. PLOS ONE, 7(1), 1-12. https://doi.org/10.1371/journal.pone.0029752 ↩
Nguyen, C., Barker, B. M., Hoover, S., Nix, D. E., Ampel, N. M., Frelinger, J. A., . . . Galgiani, J. N. (2013). Recent advances in our understanding of the environmental, epidemiological, immunological, and clinical dimensions of coccidioidomycosis. Clinical Microbiology Reviews, 26(3), 505-525. https://doi.org/10.1128/CMR.00005-13 ↩
Nicolle, L., Rotstein, C., Bourgault, A., St-Germain, G., Garber, G., & Canadian Infectious Diseases Society Invasive Fungal Registry (1998). Invasive fungal infections in Canada from 1992 to 1994. The Canadian journal of infectious diseases = Journal canadien des maladies infectieuses, 9(6), 347–352. doi:10.1155/1998/473219
↩
Nishi, J. S., Stephen, C., & Elkin, B. T. (2002). Implications of agricultural and wildlife policy on management and eradication of bovine tuberculosis and brucellosis in free-ranging wood bison of northern Canada. Annals of the New York Academy of Sciences, 969(1), 236-244. https://doi.org/10.1111/j.1749-6632.2002.tb04385.x ↩
Nyström, M.E., Karltun, J., Keller, C., Andersson Gäre B. (2018). Collaborative and partnership research for improvement of health and social services: researcher’s experiences from 20 projects. Health Research Policy and Systems, 16, 46 doi:10.1186/s12961-018-0322-0
↩
O’Brien, S. F., Delage, G., Scalia, V., Lindsay, R., Bernier, F., Dubuc, S., . . . Fearon, M. A. (2016). Seroprevalence of Babesia microti infection in Canadian blood donors. Transfusion, 56(1), 237-243. https://doi.org/10.1111/trf.13339 ↩
O’Connor, S. M., Taylor, C. E., & Hughes, J. M. (2006). Emerging infectious determinants of chronic diseases. Emerging Infectious Diseases, 12(7), 1051-1057. https://doi.org/10.3201/eid1207.060037 ↩
O’Lorcain, P. (1995). The effects of freezing on the viability of Toxocara canis and T. cati embryonated eggs. Journal of Helminthology, 69(2), 169-171. https://doi.org/10.1017/S0022149X00014073 ↩
Ogden, N. H. (2014). Lyme disease and climate change. In C. D. Butler (Ed.), Climate change and global health (pp. 145-157). Wallingford, United Kingdom: CABI.
↩
Ogden, N. H. (2017). Climate change and vector-borne diseases of public health significance. FEMS Microbiology Letters, 364(19), 1-8. https://doi.org/10.1093/femsle/fnx186 ↩
Ogden, N. H., AbdelMalik, P., & Pulliam, J. R. C. (2017). Emerging infectious diseases: Prediction and detection. Canada Communicable Disease Report, 43(10), 206-211. https://doi.org/10.14745/ccdr.v43i10a03 ↩
Ogden, N. H., Bigras-Poulin, M., O’Callaghan, C. J., Barker, I. K., Lindsay, L. R., Maarouf, A., . . . Charron, D. (2005). A dynamic population model to investigate effects of climate on geographic range and seasonality of the tick Ixodes scapularis. International Journal for Parasitology, 35(4), 375-389. https://doi.org/10.1016/j.ijpara.2004.12.013 ↩
Ogden, N. H., Bouchard, C., Kurtenbach, K., Margos, G., Lindsay, L. R., Trudel, L., . . . Milord, F. (2010). Active and passive surveillance and phylogenetic analysis of Borrelia burgdorferi elucidate the process of Lyme disease risk emergence in Canada. Environmental Health Perspectives, 118(7), 909-914. https://doi.org/10.1289/ehp.0901766 ↩
Ogden, N. H., & Gachon, P. (2019). Climate change and infectious diseases: What can we expect?. Canada communicable disease report = Releve des maladies transmissibles au Canada, 45(4), 76–80. https://doi.org/10.14745/ccdr.v45i04a01 ↩
Ogden, N. H., Koffi, J. K., Pelcat, Y., & Lindsay, L. R. (2014c). Environmental risk from Lyme disease in central and eastern Canada: A summary of recent surveillance information. Canada Communicable Disease Report, 40(5), 74-82. https://doi.org/10.14745/ccdr.v40i05a01 ↩
Ogden, N.H., Maarouf, A., Barker, I.K., Bigras-Poulin, M., Lindsay, L.R., Morshed, M.G., O’Callaghan, C.J., Ramay, F., Waltner-Toews, D. & Charron, D.F. (2006). Climate change and the potential for range expansion of the Lyme disease vector Ixodes scapularis in Canada. International Journal for Parasitology, 36, 63-70.
↩
Ogden, N. H., & Lindsay, L. R. (2016). Effects of climate and climate change on vectors and vector-borne diseases: Ticks are different. Trends in Parasitology, 32(8), 646-656. https://doi.org/10.1016/j.pt.2016.04.015 ↩
Ogden, N. H., Lindsay, L. R., Hanincová, K., Barker, I. K., Bigras-Poulin, M., Charron, D. F., . . . Thompson, R. A. (2008). Role of migratory birds in introduction and range expansion of Ixodes scapularis ticks and of Borrelia burgdorferi and Anaplasma phagocytophilum in Canada. Applied and Environmental Microbiology, 74(6), 1780-1790. https://doi.org/10.1128/AEM.01982-07 ↩
Ogden, N. H., Lindsay, L. R., & Leighton, P. A. (2013). Predicting the rate of invasion of the agent of Lyme disease, Borrelia burgdorferi. Journal of Applied Ecology, 50(2), 510-518. https://doi.org/10.1111/1365-2664.12050 ↩
Ogden, N. H., Lindsay, L. R., Ludwig, A., Morse, A. P., Zheng, H., & Zhu, H. (2019). Weather-based forecasting of mosquito-borne disease outbreaks in Canada. Canada Communicable Disease Report, 45(5), 127-132. https://doi.org/10.14745/ccdr.v45i05a03 ↩
Ogden, N. H., Lindsay, L. R., & Schofield, S. W. (2015). Methods to prevent tick bites and Lyme disease. Clinics in Laboratory Medicine, 35(4), 883-899. https://doi.org/10.1016/j.cll.2015.07.003 ↩
Ogden, N. H., Margos, G., Artsob, H., & Tsao, J. I. (2014b). Non-rickettsial tick-borne bacteria and the diseases they cause. In D. E. Sonenshine & R. M. Roe (Eds.), Biology of ticks (2nd ed., Vol. 2, pp. 278-312). New York, NY: Oxford University Press.
↩
Ogden, N. H., Pang, G., Ginsberg, H. S., Hickling, G. J., Burke, R. L., Beati, L., & Tsao, J. I. (2018). Evidence for geographic variation in life-cycle processes affecting phenology of the Lyme disease vector Ixodes scapularis (Acari: Ixodidae) in the United States. Journal of Medical Entomology, 55(6), 1386-1401. https://doi.org/10.1093/jme/tjy104 ↩
Ogden, N. H., Radojević, M., Caminade, C., & Gachon, P. (2014a). Recent and projected future climatic suitability of North America for the Asian tiger mosquito Aedes albopictus. Parasites & Vectors, 7, 1-14. https://doi.org/10.1186/s13071-014-0532-4 ↩
Ogden, N. H., Wilson, J. R. U., Richardson, D. M., Hui, C., Davies, S. J., Kumschick, S., . . . Pulliam, J. R. C. (2019). Emerging infectious diseases and biological invasions: A call for a One Health collaboration in science and management. Royal Society Open Science, 6(3), 1-15. https://doi.org/10.1098/rsos.181577 ↩
Oliveira, A. R. S., Piaggio, J., Cohnstaedt, L. W., McVey, D. S., & Cernicchiaro, N. (2018). A quantitative risk assessment (QRA) of the risk of introduction of the Japanese encephalitis virus (JEV) in the United States via infected mosquitoes transported in aircraft and cargo ships. Preventive Veterinary Medicine, 160, 1-9. https://doi.org/10.1016/j.prevetmed.2018.09.020 ↩
Onozuka, D., & Hashizume, M. (2011). The influence of temperature and humidity on the incidence of hand, foot, and mouth disease in Japan. Science of the Total Environment, 410-411, 119-125. https://doi.org/10.1016/j.scitotenv.2011.09.055 ↩
Ooi, E.-E., & Gubler, D. J. (2009). Global spread of epidemic dengue: the influence of environmental change. Future Virology, 4(6), 571-580. 10.2217/fvl.09.55
↩
O’Reilly, K. M., Auzenbergs, M., Jafari, Y., Liu, Y., Flasche, S., & Lowe, R. (2020). Effective transmission across the globe: the role of climate in COVID-19 mitigation strategies. The Lancet. Planetary health, 4(5), e172. https://doi.org/10.1016/S2542-5196(20)30106-6 ↩
Otten A., Fazil, A., Chemeris, A., Breadner, P., & Ng, V. (2019). Prioritization of vector-borne diseases in Canada under current climate and projected climate change. Microbial Risk Analysis. https://doi.org/10.1016/j.mran.2019.100089 ↩
Overy, D. P., McBurney, S., Muckle, A., Lund, L., Lewis, P. J., & Strang, R. (2016). Cryptococcus gattii VGIIb-like variant in white-tailed deer, Nova Scotia, Canada. Emerging Infectious Diseases, 22(6), 1131-1133. 10.3201/eid2206.160081
↩
Paaijmans, K. P., Cator, L. J., & Thomas, M. B. (2013). Temperature-dependent pre-bloodmeal period and temperature-driven asynchrony between parasite development and mosquito biting rate reduce malaria transmission intensity. PLOS ONE, 8(1), 1-7. 10.1371/journal.pone.0055777
↩
Paaijmans, K. P., Read, A. F., & Thomas, M. B. (2009). Understanding the link between malaria risk and climate. Proceedings of the National Academy of Sciences of the United States of America, 106(33), 13844-13849. 10.1073/pnas.0903423106
↩
Pandemic Influenza Outbreak Research Modelling Team. (2009). Modelling an influenza pandemic: a guide for the perplexed. CMAJ, 181(3-4), 171-173. 10.1503/cmaj.090885
↩
Panic, M., & Ford, J. D. (2013). A review of national-level adaptation planning with regards to the risks posed by climate change on infectious diseases in 14 OECD nations. International Journal of Environmental Research and Public Health, 10(12), 7083-7109. https://doi.org/10.3390/ijerph10127083 ↩
Papp, Z., Clark, R. G., Parmley, E. J., Leighton, F. A., Waldner, C., & Soos, C. (2017). The ecology of avian influenza viruses in wild dabbling ducks (Anas spp.) in Canada. PLOS ONE, 12(5), 1-18. https://doi.org/10.1371/journal.pone.0176297 ↩
Park, Benjamin J., Sigel, K., Vaz, V., Komatsu, K., McRill, C., Phelan, M., . . . Hajjeh, Rana A. (2005). An epidemic of coccidioidomycosis in Arizona associated with climatic changes, 1998-2001. The Journal of Infectious Diseases, 191(11), 1981-1987. https://doi.org/10.1086/430092 ↩
Parkinson, A. J., Evengard, B., Semenza, J. C., Ogden, N., Børresen, M. L., Berner, J., . . . Albihn, A. (2014). Climate change and infectious diseases in the Arctic: Establishment of a circumpolar working group. International Journal of Circumpolar Health, 73(1), 1-7. https://doi.org/10.3402/ijch.v73.25163 ↩
Pastula, D. M., Hoang Johnson, D. K., White, J. L., Dupuis, A. P., Fischer, M., & Staples, J. E. (2015). Jamestown Canyon virus disease in the United States—2000-2013. The American Journal of Tropical Medicine and Hygiene, 93(2), 384-389. https://doi.org/10.4269/ajtmh.15-0196 ↩
Patel, M. M., Pitzer, V. E., Alonso, W. J., Vera, D., Lopman, B., Tate, J., . . . Parashar, U. D. (2013). Global seasonality of rotavirus disease. The Pediatric Infectious Disease Journal, 32(4), e134-e147. https://doi.org/10.1097/INF.0b013e31827d3b68 ↩
Patz, J. A., Githeko, A. K., McCarty, J. P., Hussein, S., Confalonieri, U., & de Wet, N. (2003). Climate change and infectious diseases. In A. J. McMichael, D. H. Campbell-Lendrum, C. F. Corvalán, K. L. Ebi, A. Githeko, J. D. Scheraga, & A. Woodward (Eds.), Climate change and human health: Risks and responses (pp. 103-132). Geneva, Switzerland: World Health Organization.
↩
Patz, J.A., Campbell-Lendrum, D., Holloway, T. & Foley, J.A. (2005). Impact of regional climate change on human health. Nature, 438, 310-7. doi: 10.1038/nature04188.
↩
Paz, S. (2015). Climate change impacts on West Nile virus transmission in a global context. Philosophical Transactions of the Royal Society B: Biological Sciences, 370(1665), 1-11. https://doi.org/10.1098/rstb.2013.0561 ↩
Paz, S., & Semenza, J. C. (2016). El Niño and climate change—Contributing factors in the dispersal of Zika virus in the Americas? The Lancet, 387(10020), 745. https://doi.org/10.1016/S0140-6736(16)00256-7 ↩
Pequeno, P., Mendel, B., Rosa, C., Bosholn, M., Souza, J. L., Baccaro, F., Barbosa, R., & Magnusson, W. (2020). Air transportation, population density and temperature predict the spread of COVID-19 in Brazil. PeerJ, 8, e9322. https://doi.org/10.7717/peerj.9322 ↩
Peters, M., Godfrey, C., McInerney, P., Soares, C. B., Khalil, H., & Parker, D. (2015). The Joanna Briggs Institute reviewers’ manual 2015: Methodology for JBI scoping reviews. Adelaide, Australia: The Joanna Briggs Institute.
↩
Petersen, L. R., Brault, A. C., & Nasci, R. S. (2013a). West Nile virus: Review of the literature. JAMA, 310(3), 308-315. https://doi.org/10.1001/jama.2013.8042 ↩
Petersen, L. R., Carson, P. J., Biggerstaff, B. J., Custer, B., Borchardt, S. M., & Busch, M. P. (2013b). Estimated cumulative incidence of West Nile virus infection in US adults, 1999–2010. Epidemiology & Infection, 141(3), 591-595. https://doi.org/10.1017/s0950268812001070 ↩
Pfister, J. R., Archer, J. R., Hersil, S., Boers, T., Reed, K. D., Meece, J. K., . . . Davis, J. P. (2011). Non-rural point source blastomycosis outbreak near a yard waste collection site. Clinical Medicine & Research, 9(2), 57-65. https://doi.org/10.3121/cmr.2010.958 ↩
Phillips, P., Galanis, E., MacDougall, L., Chong, M. Y., Balshaw, R., Cook, V. J., . . . Thompson, D. (2015). Longitudinal clinical findings and outcome among patients with Cryptococcus gattii infection in British Columbia. Clinical Infectious Diseases, 60(9), 1368-1376. https://doi.org/10.1093/cid/civ041 ↩
Picard-Masson, M., Lajoie, É., Lord, J., Lalancette, C., Marchand, G., Levac, É., . . . Lajoie, L. (2016). Two related occupational cases of Legionella longbeachae infection, Quebec, Canada. Emerging Infectious Diseases, 22(7), 1289-1291. https://doi.org/10.3201/eid2207.160184 ↩
Plummer, F. A., Jones, S. M. (2017), The story of Canada’s Ebola vaccine. Canadian Medical Association Journal, 189 (43), E1326-E1327
↩
Plummer, P. J., McClure, J. T., Menzies, P., Morley, P. S., Van den Brom, R., & Van Metre, D. C. (2018). Management of Coxiella burnetii infection in livestock populations and the associated zoonotic risk: A consensus statement. Journal of Veterinary Internal Medicine, 32(5), 1481-1494. https://doi.org/10.1111/jvim.15229 ↩
Proctor, M. E., Klein, B. S., Jones, J. M., & Davis, J. P. (2002). Cluster of pulmonary blastomycosis in a rural community: Evidence for multiple high-risk environmental foci following a sustained period of diminished precipitation. Mycopathologia, 153(3), 113-120. https://doi.org/10.1023/A:1014515230994 ↩
Public Health Agency of Canada. (2009). Case definitions for communicable diseases under national surveillance. Cananda Communicable Disease Report, 35(Suppl.2).
↩
Raghavan, R. K., Peterson, A. T., Cobos, M. E., Ganta, R., & Foley, D. (2019). Current and future distribution of the lone star tick, Amblyomma americanum (L.) (Acari: Ixodidae) in North America. PLOS ONE, 14(1), 1-13. https://doi.org/10.1371/journal.pone.0209082 ↩
Rajagopalan, S., & Yoshikawa, T. T. (2016). Norovirus infections in long-term care facilities. Journal of the American Geriatrics Society, 64(5), 1097-1103. https://doi.org/10.1111/jgs.14085 ↩
Randazzo, W., Truchado, P., Cuevas-Ferrando, E., Simón, P., Allende, A., & Sánchez, G. (2020). SARS-CoV-2 RNA in wastewater anticipated COVID-19 occurrence in a low prevalence area. Water research, 181, 115942. https://doi.org/10.1016/j.watres.2020.115942 ↩
Randolph, S. E. (2004). Faune, climat et politique: Causes possibles aux récents accroissements des zoonoses à tiques [Fauna, climate and politics: Possible causes for the recent increases in tick-borne zoonoses]. Archives de Pédiatrie, 11(10), 1282-1285. https://doi.org/10.1016/j.arcped.2003.12.019 ↩
Rausch, R. (1956). Studies on the helminth fauna of Alaska: XXX. The occurrence of Echinococcus multilocularis Leuckart, 1863, on the mainland of Alaska. The American Journal of Tropical Medicine and Hygiene, 5(6), 1086-1092. https://doi.org/10.4269/ajtmh.1956.5.1086 ↩
Reed, K. D., Meece, J. K., Archer, J. R., & Peterson, A. T. (2008). Ecologic niche modeling of Blastomyces dermatitidis in Wisconsin. PLOS ONE, 3(4), 1-7. https://doi.org/10.1371/journal.pone.0002034 ↩
Reeves, W. C., Hardy, J. L., Reisen, W. K., & Milby, M. M. (1994). Potential effect of global warming on mosquito-borne arboviruses. Journal of Medical Entomology, 31(3), 323-332. https://doi.org/10.1093/jmedent/31.3.323 ↩
Reina Ortiz, M., Le, N. K., Sharma, V., Hoare, I., Quizhpe, E., Teran, E., . . . Izurieta, R. (2017). Post-earthquake Zika virus surge: Disaster and public health threat amid climatic conduciveness. Scientific Reports, 7, 1-10. https://doi.org/10.1038/s41598-017-15706-w ↩
Reisen, W., & Brault, A. C. (2007). West Nile virus in North America: Perspectives on epidemiology and intervention. Pest Management Science, 63(7), 641-646. https://doi.org/10.1002/ps.1325 ↩
Reiter, P. (2001). Climate change and mosquito-borne disease. Environmental Health Perspectives, 109(suppl 1), 141–161.
↩
Reiter, P. (2008). Climate change and mosquito-borne disease: Knowing the horse before hitching the cart. OIE Scientific and Technical Review, 27(2), 383-398. https://doi.org/10.20506/rst.27.2.1804 ↩
Revich, B., Tokarevich, N., & Parkinson, A. J. (2012). Climate change and zoonotic infections in the Russian Arctic. International Journal of Circumpolar Health, 71(1), 1-8. https://doi.org/10.3402/ijch.v71i0.18792 ↩
Rezza, G., Nicoletti, L., Angelini, R., Romi, R., Finarelli, A. C., Panning, M., . . . Cassone, A. (2007). Infection with chikungunya virus in Italy: An outbreak in a temperate region. The Lancet, 370(9602), 1840-1846. https://doi.org/10.1016/s0140-6736(07)61779-6 ↩
Ripoche, M., Campagna, C., Ludwig, A., Ogden, N. H., & Leighton, P. A. (2019). Short-term forecasting of daily abundance of West Nile virus vectors Culex pipiens-restuans (Diptera: Culicidae) and Aedes vexans based on weather conditions in southern Québec (Canada). Journal of Medical Entomology, 56(3), 859-872. https://doi.org/10.1093/jme/tjz002 ↩
Roberts, K. C., Rao, D. P., Bennett, T. L., Loukine, L., Jayaraman, G. C. (2015). Prevalence and patterns of chronic disease multimorbidity and associated determinants in Canada. Health Promotion and Chronic Disease Prevention Canada, 35(6), 87-94. doi: 10.24095/hpcdp.35.6.01. PMID: 26302227; PMCID: PMC4910465
↩
Rocheleau, J. P., Michel, P., Lindsay, L. R., Drebot, M., Dibernardo, A., Ogden, N. H., . . . Arsenault, J. (2018). Risk factors associated with seropositivity to California serogroup viruses in humans and pet dogs, Quebec, Canada. Epidemiology & Infection, 146(9), 1167-1176. https://doi.org/10.1017/s0950268818001000 ↩
Rodenhouse, N. L., Christenson, L. M., Parry, D., & Green, L. E. (2009). Climate change effects on native fauna of northeastern forests. Canadian Journal of Forest Research, 39(2), 249-263. https://doi.org/10.1139/X08-160 ↩
Roe, C. C., Bowers, J., Oltean, H., DeBess, E., Dufresne, P. J., McBurney, S., . . . Engelthaler, D. M. (2018). Dating the Cryptococcus gattii dispersal to the North American Pacific Northwest. mSphere, 3(1), 1-13. https://doi.org/10.1128/mSphere.00499-17 ↩
Rohr, J. R., Dobson, A. P., Johnson, P. T. J., Kilpatrick, A. M., Paull, S. H., Raffel, T. R., . . . Thomas, M. B. (2011). Frontiers in climate change-disease research. Trends in Ecology & Evolution, 26(6), 270-277. https://doi.org/10.1016/j.tree.2011.03.002 ↩
Romeo-Aznar, V., Paul, R., Telle, O., & Pascual, M. (2018). Mosquito-borne transmission in urban landscapes: The missing link between vector abundance and human density. Proceedings of the Royal Society B: Biological Sciences, 285(1884), 1-9. https://doi.org/10.1098/rspb.2018.0826 ↩
Romero-Lankao, P., Smith, J. B., Davidson, D. J., Diffenbaugh, N. S., Kinney, P. L., Kirshen, P., . . . Villers Ruiz, L. (2014). North America. In V. R. Barros, C. B. Field, D. J. Dokken, M. D. Mastrandrea, K. J. Mach, T. E. Bilir, M. Chatterjee, K. L. Ebi, Y. O. Estrada, R. C. Genova, B. Girma, E. S. Kissel, A. N. Levy, S. MacCracken, P. R. Mastrandrea, & L. L. White (Eds.), Climate change 2014: Impacts, adaptation, and vulnerability. Part B: Regional aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (pp. 1439-1498). Cambridge, England: Cambridge University Press. Retrieved from https://www.ipcc.ch/report/ar5/wg2/ ↩
Rosas, Á. L., & Casadevall, A. (2006). Melanization affects susceptibility of Cryptococcus neoformans to heat and cold. FEMS Microbiology Letters, 153(2), 265-272. https://doi.org/10.1111/j.1574-6968.1997.tb12584.x ↩
Rosenstein, N. E., Perkins, B. A., Stephens, D. S., Popovic, T., & Hughes, J. M. (2001). Meningococcal disease. The New England Journal of Medicine, 344(18), 1378-1388. https://doi.org/10.1056/NEJM200105033441807 ↩
Roussel, M., Pontier, D., Cohen, J.-M., Lina, B., & Fouchet, D. (2016). Quantifying the role of weather on seasonal influenza. BMC Public Health, 16, 1-14. https://doi.org/10.1186/s12889-016-3114-x ↩
Rycroft-Malone, J., Burton, C., Wilkinson, J., Harvey, G., McCormack, B., Baker, R., Dopson, S., Graham, I.D., Staniszewska, S., Thompson, C., Ariss, S., Melville-Richards, L., Williams, L. (2015). Collective action for implementation: a realist evaluation of organisational collaboration in healthcare. Implementation Science, 11(17). doi:10.1186/s13012-016-0380-z
↩
Safronetz, D., Sloan, A., Stein, D. R., Mendoza, E., Barairo, N., Ranadheera, C., . . . Drebot, M. (2017). Evaluation of 5 commercially available Zika virus immunoassays. Emerging Infectious Diseases, 23(9), 1577-1580. https://doi.org/10.3201/eid2309.162043 ↩
Sage, K. M., Johnson, T. L., Teglas, M. B., Nieto, N. C., & Schwan, T. G. (2017). Ecological niche modeling and distribution of Ornithodoros hermsi associated with tick-borne relapsing fever in western North America. PLOS Neglected Tropical Diseases, 11(10), 1-18. https://doi.org/10.1371/journal.pntd.0006047 ↩
Sagurova I, Ludwig A, Ogden NH, Pelcat Y, Dueymes G, Gachon P. (2019). Predicted northward expansion of the geographic range of the tick vector Amblyomma americanum in North America under future climate conditions. Environmental Health Perspectives, 127(10), 107014. doi: 10.1289/EHP5668
↩
Sakamoto, R., Ohno, A., Nakahara, T., Satomura, K., Iwanaga, S., Kouyama, Y., . . . Yamaguchi, K. (2009). Legionella pneumophila in rainwater on roads. Emerging Infectious Diseases, 15(8), 1295-1297. https://doi.org/10.3201/eid1508.090317 ↩
Salb, A. L., Barkema, H. W., Elkin, B. T., Thompson, R. C. A., Whiteside, D. P., Black, S. R., . . . Kutz, S. J. (2008). Dogs as sources and sentinels of parasites in humans and wildlife, northern Canada. Emerging Infectious Diseases, 14(1), 60-63. https://doi.org/10.3201/eid1401.071113 ↩
Salkeld DJ, Padgett KA, Jones JH. (2013). A meta-analysis suggesting that the relationship between biodiversity and risk of zoonotic pathogen transmission is idiosyncratic. Ecology Letters, 16(5), 679-686
↩
Samia, N. I., Kausrud, K. L., Heesterbeek, H., Ageyev, V., Begon, M., Chan, K. S., Stenseth, N. C. (2011) Dynamics of the plague-wildlife-human system in Central Asia are controlled by two epidemiological thresholds. Proceedings of the National Academies of Science of the U S A, 108(35), 14527-14532.
↩
Samy, A. M., Elaagip, A. H., Kenawy, M. A., Ayres, C. F., Peterson, A. T., & Soliman, D. E. (2016). Climate change influences on the global potential distribution of the mosquito Culex quinquefasciatus, vector of West Nile virus and lymphatic filariasis. PLOS ONE, 11(10), 1-13. https://doi.org/10.1371/journal.pone.0163863 ↩
Sapp, S. G. H., Rascoe, L. N., Wilkins, P. P., Handali, S., Gray, E. B., Eberhard, M. L., . . . Yabsley, M. J. (2016). Baylisascaris procyonis roundworm seroprevalence among wildlife rehabilitators, United States and Canada, 2012–2015. Emerging Infectious Diseases, 22(12), 2128-2131. https://doi.org/10.3201/eid2212.160467 ↩
Schanzer, D. L., Langley, J. M., & Tam, T. W. S. (2006). Hospitalization attributable to influenza and other viral respiratory illnesses in Canadian children. The Pediatric Infectious Disease Journal, 25(9), 795-800. https://doi.org/10.1097/01.inf.0000232632.86800.8c ↩
Schanzer, D. L., Langley, J. M., & Tam, T. W. S. (2008). Role of influenza and other respiratory viruses in admissions of adults to Canadian hospitals. Influenza and Other Respiratory Viruses, 2(1), 1-8. https://doi.org/10.1111/j.1750-2659.2008.00035.x ↩
Schanzer, D. L., McGeer, A., & Morris, K. (2013a). Statistical estimates of respiratory admissions attributable to seasonal and pandemic influenza for Canada. Influenza and Other Respiratory Viruses, 7(5), 799-808. https://doi.org/10.1111/irv.12011 ↩
Schanzer, D. L., Sevenhuysen, C., Winchester, B., & Mersereau, T. (2013b). Estimating influenza deaths in Canada, 1992-2009. PLOS ONE, 8(11), 1-11. https://doi.org/10.1371/journal.pone.0080481 ↩
Schneeberger, P. M., Wintenberger, C., van der Hoek, W., & Stahl, J. P. (2014). Q fever in the Netherlands 2007-2010: What we learned from the largest outbreak ever. Médecine et Maladies Infectieuses, 44(8), 339-353. https://doi.org/10.1016/j.medmal.2014.02.006 ↩
Schultz-Cherry, S., Olsen, C. W., & Easterday, B. C. (2011). History of swine influenza. Current Topics in Microbiology and Immunology, 370, 21-28. https://doi.org/10.1007/82_2011_197 ↩
Seitz, A. E., Adjemian, J., Steiner, C. A., & Prevots, D. R. (2015). Spatial epidemiology of blastomycosis hospitalizations: Detecting clusters and identifying environmental risk factors. Medical Mycology, 53(5), 447-454. https://doi.org/10.1093/mmy/myv014 ↩
Sejvar, J. J. (2014). Clinical manifestations and outcomes of West Nile virus infection. Viruses, 6(2), 606-623. https://doi.org/10.3390/v6020606 ↩
Semenza, J. C., Lindgren, E., Balkanyi, L., Espinosa, L., Almqvist, M. S., Penttinen, P., & Rocklöv, J. (2016). Determinants and drivers of infectious disease threat events in Europe. Emerging Infectious Diseases, 22(4), 581-589. https://doi.org/10.3201/eid2204 ↩
Septfons, A., Leparc-Goffart, I., Couturier, E., Franke, F., Deniau, J., Balestier, A., . . . De Valk, H. (2016). Travel-associated and autochthonous Zika virus infection in mainland France, 1 January to 15 July 2016. Eurosurveillance, 21(32), 1-7. https://doi.org/10.2807/1560-7917.Es.2016.21.32.30315 ↩
Shafir, S. C., Sorvillo, F. J., Sorvillo, T., & Eberhard, M. L. (2011). Viability of Baylisascaris procyonis eggs. Emerging Infectious Diseases, 17(7), 1293-1295. https://doi.org/10.3201/eid1707.101774 ↩
Shaman, J., Day, J. F., & Stieglitz, M. (2005). Drought-induced amplification and epidemic transmission of West Nile virus in southern Florida. Journal of Medical Entomology, 42(2), 134-141. https://doi.org/10.1093/jmedent/42.2.134 ↩
Shaman, J., Pitzer, V. E., Viboud, C., Grenfell, B. T., & Lipsitch, M. (2010). Absolute humidity and the seasonal onset of influenza in the continental United States. PLOS Biology, 8(2), 1-13. https://doi.org/10.1371/journal.pbio.1000316 ↩
Shapiro, L. L. M., Whitehead, S. A., & Thomas, M. B. (2017). Quantifying the effects of temperature on mosquito and parasite traits that determine the transmission potential of human malaria. PLOS Biology, 15(10), 1-21. https://doi.org/10.1371/journal.pbio.2003489 ↩
Silk, B. J., Foltz, J. L., Ngamsnga, K., Brown, E., Muñoz, M. G., Hampton, L. M., . . . Hicks, L. A. (2013). Legionnaires’ disease case-finding algorithm, attack rates, and risk factors during a residential outbreak among older adults: An environmental and cohort study. BMC Infectious Diseases, 13, 1-8. https://doi.org/10.1186/1471-2334-13-291 ↩
Simmering, J. E., Polgreen, L. A., Hornick, D. B., Sewell, D. K., & Polgreen, P. M. (2017). Weather-dependent risk for Legionnaires’ disease, United States. Emerging Infectious Diseases, 23(11), 1843-1851. https://doi.org/10.3201/eid2311.170137 ↩
Simon, A., Tardy, O., Hurford, A., Lecomte, N., Bélanger, D., & Leighton, P. (2019). Dynamics and persistence of rabies in the Arctic. Polar Research, 38, 1-14.
↩
Simon, J. A., Marrotte, R. R., Desrosiers, N., Fiset, J., Gaitan, J., Gonzalez, A., . . . Millien, V. (2014). Climate change and habitat fragmentation drive the occurrence of Borrelia burgdorferi, the agent of Lyme disease, at the northeastern limit of its distribution. Evolutionary Applications, 7(7), 750-764. https://doi.org/10.1111/eva.12165 ↩
Sivabalan, P., Saboo, A., Yew, J., & Norton, R. (2017). Q fever in an endemic region of North Queensland, Australia: A 10 year review. One Health, 3, 51-55. https://doi.org/10.1016/J.ONEHLT.2017.03.002 ↩
Skog, L., Linde, A., Palmgren, H., Hauska, H., & Elgh, F. (2014). Spatiotemporal characteristics of pandemic influenza. BMC Infectious Diseases, 14, 1-14. https://doi.org/10.1186/1471-2334-14-378 ↩
Skowronski, D. M., Li, Y., Tweed, S. A., Tam, T. W. S., Petric, M., David, S. T., . . . Brunham, R. C. (2007). Protective measures and human antibody response during an avian influenza H7N3 outbreak in poultry in British Columbia, Canada. CMAJ, 176(1), 47-53. https://doi.org/10.1503/cmaj.060204 ↩
Smith, K. R., Woodward, A., Campbell-Lendrum, D., Chadee, D. D., Honda, Y., Liu, Q., . . . Sauerborn, R. (2014). Human health: Impacts, adaptation, and co-benefits. In C. B. Field, V. R. Barros, D. J. Dokken, K. J. Mach, M. D. Mastrandrea, T. E. Bilir, M. Chatterjee, K. L. Ebi, Y. O. Estrada, R. C. Genova, B. Girma, E. S. Kissel, A. N. Levy, S. MacCracken, P. R. Mastrandrea, & L. L. White (Eds.), Climate change 2014: Impacts, adaptation, and vulnerability. Part A: Global and sectoral aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (pp. 709-754). Cambridge, England: Cambridge University Press. Retrieved from https://www.ipcc.ch/report/ar5/wg2/ ↩
Sonenshine, D. E. (2018). Range expansion of tick disease vectors in North America: Implications for spread of tick-borne disease. International Journal of Environmental Research and Public Health, 15(3), 1-9. https://doi.org/10.3390/ijerph15030478 ↩
Souto, A. C. P., Bonfietti, L. X., Ferreira-Paim, K., Trilles, L., Martins, M., Ribeiro-Alves, M., . . . Meyer, W. (2016). Population genetic analysis reveals a high genetic diversity in the Brazilian Cryptococcus gattii VGII population and shifts the global origin from the Amazon rainforest to the semi-arid desert in the northeast of Brazil. PLOS Neglected Tropical Diseases, 10(8), 1-19. https://doi.org/10.1371/journal.pntd.0004885 ↩
Spence, L., Artsob, H., Grant, L., & Th’Ng, C. (1977). St. Louis encephalitis in southern Ontario: Laboratory studies for arboviruses. CMAJ, 116(1), 35-37. Retrieved from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1879132/ ↩
St-Germain, G., Murray, G., & Duperval, R. (1993). Blastomycosis in Quebec (1981-90): Report of 23 cases and review of published cases from Quebec. Canadian Journal of Infectious Diseases, 4(2), 89-94. https://doi.org/10.1155/1993/249823 ↩
St-Pierre, J., Dufresne, P. J., Carignan, A., Lévesque, É., Bernard, F., Longtin, J., & LeBlanc, L. (2018). Case series: Report of the first two human indigenous cases of Cryptococcus gattii infection in eastern Canada. Mycopathologia, 183(2), 399-406. https://doi.org/10.1007/s11046-017-0215-8 ↩
Stafford, K. C., III., Molaei, G., Little, E. A. H., Paddock, C. D., Karpathy, S. E., & Labonte, A. M. (2018). Distribution and establishment of the lone star tick in Connecticut and implications for range expansion and public health. Journal of Medical Entomology, 55(6), 1561-1568. https://doi.org/10.1093/jme/tjy115 ↩
Sultan, B., Labadi, K., Guégan, J.-F., & Janicot, S. (2005). Climate drives the meningitis epidemics onset in West Africa. PLOS Medicine, 2(1), 0043-0049. https://doi.org/10.1371/journal.pmed.0020006 ↩
Sumi, A., Toyoda, S., Kanou, K., Fujimoto, T., Mise, K., Kohei, Y., . . . Kobayashi, N. (2017). Association between meteorological factors and reported cases of hand, foot, and mouth disease from 2000 to 2015 in Japan. Epidemiology & Infection, 145(14), 2896-2911. https://doi.org/10.1017/S0950268817001820 ↩
Sunenshine, R. H., Anderson, S., Erhart, L., Vossbrink, A., Kelly, P. C., Engelthaler, D., & Komatsu, K. (2007). Public health surveillance for coccidioidomycosis in Arizona. Annals of the New York Academy of Sciences, 1111(1), 96-102. https://doi.org/10.1196/annals.1406.045 ↩
Suresh, S., Forgie, S., & Robinson, J. (2018). Non-polio Enterovirus detection with acute flaccid paralysis: A systematic review. Journal of Medical Virology, 90(1), 3-7. https://doi.org/10.1002/jmv.24933 ↩
Tabachnick, W. J. (2016). Climate change and the arboviruses: Lessons from the evolution of the dengue and yellow fever viruses. Annual Review of Virology, 3, 125-145. https://doi.org/10.1146/annurev-virology-110615-035630 ↩
Takeuchi-Storm, N., Woolsey, I. D., Jensen, P. M., Fredensborg, B. L., Pipper, C. B., & Kapel, C. M. O. (2015). Predictors of Echinococcus multilocularis prevalence in definitive and intermediate hosts: A meta-analysis approach. Journal of Parasitology, 101(3), 297-303. https://doi.org/10.1645/14-645.1 ↩
Tam, B. Y., Martin, I., & Tsuji, L. J. S. (2014). Geospatial analysis between the environment and past incidences of West Nile virus in bird specimens in Ontario, Canada. GeoJournal, 79(6), 805-817. https://doi.org/10.1007/s10708-014-9526-1 ↩
Tamerius, J. D., & Comrie, A. C. (2011). Coccidioidomycosis incidence in Arizona predicted by seasonal precipitation. PLOS ONE, 6(6), 1-7. https://doi.org/10.1371/journal.pone.0021009 ↩
Tang, J. W., Lai, F. Y. L., Nymadawa, P., Deng, Y. M., Ratnamohan, M., Petric, M., . . . Wong, F. Y. W. (2010). Comparison of the incidence of influenza in relation to climate factors during 2000–2007 in five countries. Journal of Medical Virology, 82(11), 1958-1965. https://doi.org/10.1002/jmv.21892 ↩
Tasanee, A., Chayanit, L., & Agsornpien, N. (2015). Does the correlation between rainfall vs dengue hemorrhagic fever incidence always exist? Case study in Phuket, Thailand. Journal of Environmental Research and Development, 9(3), 509-513. Retrieved from http://jerad.org/archiveabstract.php?vol=9&issue=3 ↩
Tatem, A. J., Hay, S. I., & Rogers, D. J. (2006). Global traffic and disease vector dispersal. Proceedings of the National Academy of Sciences of the United States of America, 103(16), 6242-6247. https://doi.org/10.1073/pnas.0508391103 ↩
Tatem, A. J., Huang, Z., Das, A., Qi, Q., Roth, J., & Qiu, Y. (2012). Air travel and vector-borne disease movement. Parasitology, 139(14), 1816-1830. https://doi.org/10.1017/S0031182012000352 ↩
Teixeira, M. de M., Patan, J. S., Taylor, M. L., Gómez, B. L., Theodoro, R. C., de Hoog, S., Engelthaler, D. M., Zancopé-Oliveira, R. M., Felipe, M. S., Barker, B. M. (2016). Worldwide phylogenetic distributions and population dynamics of the genus Histoplasma. PLoS Neglected Tropical Diseases 10, e0004732. doi: 10.1371/journal.pntd.0004732.
↩
Thielman, A. C., & Hunter, F. F. (2007). Photographic key to the adult female mosquitoes (Diptera: Culicidae) of Canada. Canadian Journal of Arthropod Identification, 04. https://doi.org/10.3752/cjai.2007.04 ↩
Thomas-Bachli, A. L., Pearl, D. L., Berke, O., Parmley, E. J., & Barker, I. K. (2015). A comparison of West Nile virus surveillance using survival analyses of dead corvid and mosquito pool data in Ontario, 2002–2008. Preventive Veterinary Medicine, 122(3), 363-370. https://doi.org/10.1016/j.prevetmed.2015.10.007 ↩
Thompson, W. W., Comanor, L., & Shay, D. K. (2006). Epidemiology of seasonal influenza: Use of surveillance data and statistical models to estimate the burden of disease. The Journal of Infectious Diseases, 194(Suppl. 2), S82-S91. https://doi.org/10.1086/507558 ↩
Tinline, R., & Gregory, D. (i2020). Taking the bite out of rabies: The evolution of rabies management in Canada. Toronto, ON: University of Toronto Press.
↩
Tomaselli, M., Kutz, S., Gerlach, C., & Checkley, S. (2017). Local knowledge to enhance wildlife population health surveillance: Conserving muskoxen and caribou in the Canadian Arctic. Biological Conservation, 217, 337-48.
↩
Tomasello, D., & Schlagenhauf, P. (2013). Chikungunya and dengue autochthonous cases in Europe, 2007-2012. Travel Medicine and Infectious Disease, 11(5), 274-284. https://doi.org/10.1016/j.tmaid.2013.07.006 ↩
Tong, D. Q., Wang, J. X. L., Gill, T. E., Lei, H., & Wang, B. (2017). Intensified dust storm activity and valley fever infection in the southwestern United States. Geophysical Research Letters, 44(9), 4304-4312. https://doi.org/10.1002/2017GL073524 ↩
Tossa, P., Deloge-Abarkan, M., Zmirou-Navier, D., Hartemann, P., & Mathieu, L. (2006). Pontiac fever: An operational definition for epidemiological studies. BMC Public Health, 6, 112. 10.1186/1471-2458-6-112
↩
Towers, S., Chowell, G., Hameed, R., Jastrebski, M., Khan, M., Meeks, J., . . . Harris, G. (2013). Climate change and influenza: The likelihood of early and severe influenza seasons following warmer than average winters. PLOS Currents: Influenza, 1. https://doi.org/10.1371/currents.flu.3679b56a3a5313dc7c043fb944c6f138 ↩
Tricco, A. C., Lillie, E., Zarin, W., O’Brien, K., Colquhoun, H., Kastner, M., . . . Straus, S. E. (2016). A scoping review on the conduct and reporting of scoping reviews. BMC Medical Research Methodology, 16, 1-10. https://doi.org/10.1186/s12874-016-0116-4 ↩
Trudel, L., Veillette, M., Bonifait, L., & Duchaine, C. (2014). Management of the 2012 Legionella crisis in Quebec City: Need for a better communication between resources and knowledge transfer. Frontiers in Microbiology, 5, 1-4. https://doi.org/10.3389/fmicb.2014.00182 ↩
Tsetsarkin, K. A., Vanlandingham, D. L., McGee, C. E., & Higgs, S. (2007). A single mutation in chikungunya virus affects vector specificity and epidemic potential. PLOS Pathogens, 3(12), 1895-1906. https://doi.org/10.1371/journal.ppat.0030201 ↩
Tuite, A. R., Kinlin, L. M., Kuster, S. P., Jamieson, F., Kwong, J. C., McGeer, A., & Fisman, D. N. (2010). Respiratory virus infection and risk of invasive meningococcal disease in central Ontario, Canada. PLOS ONE, 5(11), 1-7. https://doi.org/10.1371/journal.pone.0015493 ↩
Turell, M. J., Britch, S. C., Aldridge, R. L., Kline, D. L., Boohene, C., & Linthicum, K. J. (2013). Potential for mosquitoes (Diptera: Culicidae) from Florida to transmit Rift Valley fever virus. Journal of Medical Entomology, 50(5), 1111-1117. https://doi.org/10.1603/ME13049 ↩
Turell, M. J., Byrd, B. D., & Harrison, B. A. (2013). Potential for populations of Aedes j. japonicus to transmit Rift Valley fever virus in the USA. Journal of the American Mosquito Control Association, 29(2), 133-137. https://doi.org/10.2987/12-6316r.1 ↩
Turell, M. J., Dohm, D. J., & Fonseca, D. M. (2014). Comparison of the potential for different genetic forms in the Culex pipiens complex in North America to transmit Rift Valley fever virus. Journal of the American Mosquito Control Association, 30(4), 253-259. https://doi.org/10.2987/14-6441R.1 ↩
Turell, M. J., Wilson, W. C., & Bennett, K. E. (2010). Potential for North American mosquitoes (Diptera: Culicidae) to transmit Rift Valley fever virus. Journal of Medical Entomology, 47(5), 884-889. https://doi.org/10.1603/ME10007 ↩
Uejio, C. K., Mak, S., Manangan, A., Luber, G., & Bartlett, K. H. (2015). Climatic influences on Cryptococcus gattii populations, Vancouver Island, Canada, 2002-2004. Emerging Infectious Diseases, 21(11), 1989-1996. https://doi.org/10.3201/eid2111.141161 ↩
Valseth, K., Nesbø, C. L., Easterday, W. R., Turner, W. C., Olsen, J. S., Stenseth, N. C., & Haverkamp, T. H. A. (2017). Temporal dynamics in microbial soil communities at anthrax carcass sites. BMC Microbiology, 17, 1-15. https://doi.org/10.1186/s12866-017-1111-6 ↩
van Leuken, J. P. G., Swart, A. N., Droogers, P., van Pul, A., Heederik, D., & Havelaar, A. H. (2016). Climate change effects on airborne pathogenic bioaerosol concentrations: A scenario analysis. Aerobiologia, 32(4), 607-617. https://doi.org/10.1007/s10453-016-9435-5 ↩
Vander Kelen, P., Downs, J. A., Unnasch, T., & Stark, L. (2014). A risk index model for predicting eastern equine encephalitis virus transmission to horses in Florida. Applied Geography, 48, 79-86. https://doi.org/10.1016/j.apgeog.2014.01.012 ↩
Varia, M., Wilson, S., Sarwal, S., McGeer, A., Gournis, E., Galanis, E., & Henry, B. (2003). Investigation of a nosocomial outbreak of severe acute respiratory syndrome (SARS) in Toronto, Canada. CMAJ, 169(4), 285-292. Retrieved from http://www.cmaj.ca/content/169/4/285.long ↩
Varrin, R., Bowman, J., & Gray, P. A. (2007). The known and potential impacts of climate change on biodiversity in Ontario’s terrestrial ecosystems: Case studies and recommendations for adaptation. (Climate Change Research Report CCRR-09). Sault Ste. Marie, ON: Ontario Ministry of Natural Resources Retrieved from http://www.climateontario.ca/MNR_Publications/276927.pdf ↩
Venturi, G., Di Luca, M., Fortuna, C., Remoli, M. E., Riccardo, F., Severini, F., . . . Rizzo, C. (2017). Detection of a chikungunya outbreak in Central Italy, August to September 2017. Eurosurveillance, 22(39), 1-4. https://doi.org/10.2807/1560-7917.ES.2017.22.39.17-00646 ↩
Viboud, C., Pakdaman, K., Boëlle, P.-Y., Wilson, M. L., Myers, M. F., Valleron, A.-J., & Flahault, A. (2004). Association of influenza epidemics with global climate variability. European Journal of Epidemiology, 19(11), 1055-1059. https://doi.org/10.1007/s10654-004-2450-9 ↩
Villeneuve, A., Polley, L., Jenkins, E., Schurer, J., Gilleard, J., Kutz, S., . . . Gagné, F. (2015). Parasite prevalence in fecal samples from shelter dogs and cats across the Canadian provinces. Parasites & Vectors, 8, 1-10. https://doi.org/10.1186/s13071-015-0870-x ↩
Vugia, D. J., Wheeler, C., Cummings, K. C., & Karon, A. (2009). Increase in coccidioidomycosis—California, 2000-2007. MMWR Morbidity and Mortality Weekly Report, 58(5), 105-109. Retrieved from https://www.cdc.gov/mmwr/preview/mmwrhtml/mm5805a1.htm ↩
Walsh, M. G., de Smalen, A. W., & Mor, S. M. (2018). Climatic influence on anthrax suitability in warming northern latitudes. Scientific Reports, 8, 1-9. https://doi.org/10.1038/s41598-018-27604-w ↩
Wang, C., Liu, L., Hao, X., Guo, H., Wang, Q., Huang, J., He, N., Yu, H., Lin, X., Pan, A., Wei, S., Wu, T. (2020). Evolving Epidemiology and Impact of Non-pharmaceutical Interventions on the Outbreak of Coronavirus Disease 2019 in Wuhan, China. JAMA. doi:10.1101/2020.03.03.20030593
↩
Wang, G., Minnis, R. B., Belant, J. L., & Wax, C. L. (2010). Dry weather induces outbreaks of human West Nile virus infections. BMC Infectious Diseases, 10, 1-7. https://doi.org/10.1186/1471-2334-10-38 ↩
Wang, J., Ogden, N. H., & Zhu, H. (2011). The impact of weather conditions on Culex pipiens and Culex restuans (Diptera: Culicidae) abundance: A case study in Peel Region. Journal of Medical Entomology, 48(2), 468-475. https://doi.org/10.1603/ME10117 ↩
Wang, P., Goggins, W. B., & Chan, E. Y. Y. (2016). Hand, foot and mouth disease in Hong Kong: A time-series analysis on its relationship with weather. PLOS ONE, 11(8), 1-12. https://doi.org/10.1371/journal.pone.0161006 ↩
Webster, D., Dimitrova, K., Holloway, K., Makowski, K., Safronetz, D., & Drebot, M. A. (2017). California serogroup virus infection associated with encephalitis and cognitive decline, Canada, 2015. Emerging Infectious Diseases, 23(8), 1423-1424. https://doi.org/10.3201/eid2308.170239 ↩
Weissenböck, H., Kolodziejek, J., Url, A., Lussy, H., Rebel-Bauder, B., & Nowotny, N. (2002). Emergence of Usutu virus, an African mosquito-borne Flavivirus of the Japanese encephalitis virus group, central Europe. Emerging Infectious Diseases, 8(7), 652-656. https://doi.org/10.3201/eid0807.020094 ↩
White, A. N. J., Ng, V., Spain, C. V., Johnson, C. C., Kinlin, L. M., & Fisman, D. N. (2009). Let the sun shine in: Effects of ultraviolet radiation on invasive pneumococcal disease risk in Philadelphia, Pennsylvania. BMC Infectious Diseases, 9. https://doi.org/10.1186/1471-2334-9-196 ↩
Williams, C. R., Mincham, G., Faddy, H., Viennet, E., Ritchie, S. A., & Harley, D. (2016). Projections of increased and decreased dengue incidence under climate change. Epidemiology & Infection, 144(14), 3091-3100. https://doi.org/10.1017/s095026881600162x ↩
Wilson, W. C., Kim, I. J., Trujillo, J. D., Sunwoo, S. Y., Noronha, L. E., Urbaniak, K., . . . Richt, J. A. (2018). Susceptibility of white-tailed deer to Rift Valley fever virus. Emerging Infectious Diseases, 24(9), 1717-1719. https://doi.org/10.3201/eid2409.180265 ↩
Wong, S.S.Y., Yip, C.C.Y., Lau, S.K.P. & Yuen, K.Y. (2010). Human enterovirus 71 and hand, foot and mouth disease. Epidemiology & Infection, 13, 1071-89. doi: 10.1017/S0950268809991555.
↩
Wood, D. M., Dang, P. T., & Ellis, R. A. (1979). Part 6 The mosquitoes of Canada—Diptera: Culicidae. In The insects and arachnids of Canada. Ottawa, ON: Government of Canada. Retrieved from http://publications.gc.ca/pub?id=9.811351&sl=0 ↩
World Health Organization. (2013). WHO expert consultation on rabies: Second report (WHO Technical Report Series 982). Geneva, Switzerland: WHO Press. Retrieved from https://apps.who.int/iris/handle/10665/85346 ↩
World Health Organization (WHO). (2019c). Strengthening health security by implementing the International Health Regulations (2005). Retrieved from https://www.who.int/ihr/en/ ↩
World Tourism Organization, & United Nations Environment Programme. (Eds.). (2008). Climate change and tourism—Responding to global challenges. Madrid, Spain: World Tourism Organization.
↩
Wormser, G. P., Dattwyler, R. J., Shapiro, E. D., Halperin, J. J., Steere, A. C., Klempner, M. S., . . . Nadelman, R. B. (2006). The clinical assessment, treatment, and prevention of Lyme disease, human granulocytic anaplasmosis, and babesiosis: Clinical practice guidelines by the Infectious Diseases Society of America. Clinical Infectious Diseases, 43(9), 1089-1134. https://doi.org/10.1086/508667 ↩
Wu, J. T., Leung, K., & Leung, G. M. (2020). Nowcasting and forecasting the potential domestic and international spread of the 2019-nCoV outbreak originating in Wuhan, China: A modelling study. The Lancet,395(10225), 689-697. doi:10.1016/s0140-6736(20)30260-9
↩
Xiao, F.-Z., Zhang, Y., Deng, Y.-Q., He, S., Xie, H.-G., Zhou, X.-N., & Yan, Y.-S. (2014). The effect of temperature on the extrinsic incubation period and infection rate of dengue virus serotype 2 infection in Aedes albopictus. Archives of Virology, 159(11), 3053-3057. https://doi.org/10.1007/s00705-014-2051-1 ↩
Yang, H. M., Macoris, M. L. G., Galvani, K. C., Andrighetti, M. T. M., & Wanderley, D. M. V. (2009). Assessing the effects of temperature on the population of Aedes aegypti, the vector of dengue. Epidemiology & Infection, 137(8), 1188-1202. https://doi.org/10.1017/s0950268809002040 ↩
Yansouni, C.P., Pernica, J.M., & Goldfarb, D. (2016). Enteric parasites in Arctic communities: Tip of the iceberg? Trends in Parasitology, 32(11), 834-38.
↩
Yoo, E. H., Chen, D., Diao, C., & Russell, C. (2016). The effects of weather and environmental factors on West Nile virus mosquito abundance in Greater Toronto Area. Earth Interactions, 20(3), 1-22. https://doi.org/10.1175/ei-d-15-0003.1 ↩
Yu, G., Li, Y., Cai, J., Yu, D., Tang, J., Zhai, W., . . . Qin, J. (2019). Short-term effects of meteorological factors and air pollution on childhood hand-foot-mouth disease in Guilin, China. Science of the Total Environment, 646, 460-470. https://doi.org/10.1016/j.scitotenv.2018.07.329 ↩
Yue, R. P. H., & Lee, H. F. (2018). Pre-industrial plague transmission is mediated by the synergistic effect of temperature and aridity index. BMC Infectious Diseases, 18(1), 134. 10.1186/s12879-018-3045-5
↩
Zhang, T., Wu, Q., & Zhang, Z. (2020). Probable Pangolin Origin of SARS-CoV-2 Associated with the COVID-19 Outbreak. Current biology : CB, 30(8), 1578. https://doi.org/10.1016/j.cub.2020.03.063 ↩
Zhao, D., Wang, L., Cheng, J., Xu, J., Xu, Z., Xie, M., . . . Su, H. (2017). Impact of weather factors on hand, foot and mouth disease, and its role in short-term incidence trend forecast in Huainan City, Anhui province. International Journal of Biometeorology, 61(3), 453-461. https://doi.org/10.1007/s00484-016-1225-9 ↩
Zhao, Q., Li, S., Cao, W., Liu, D.-L., Qian, Q., Ren, H., . . . Guo, Y. (2018). Modeling the present and future incidence of pediatric hand, foot, and mouth disease associated with ambient temperature in mainland China. Environmental Health Perspectives, 126(4), 1-11. https://doi.org/10.1289/EHP3062 ↩
Zheng, H., Drebot, M. A., & Coulthart, M. B. (2014). West Nile virus in Canada: Ever-changing, but here to stay. Canada Communicable Disease Report, 40(10), 173-177. https://doi.org/10.14745/ccdr.v40i10a01 ↩
Zhu, F., Xu, W., Xia, J., Liang, Z., Liu, Y., Zhang, X., . . . Wang, N. (2014). Efficacy, safety, and immunogenicity of an enterovirus 71 vaccine in China. The New England Journal of Medicine, 370(9), 818-828. https://doi.org/10.1056/NEJMoa1304923 ↩